






肺癌治疗的手段-AIF
凋亡诱导因子(AIF)不仅参与细胞凋亡,还在线粒体内发挥重要的功能,它通过线粒体呼吸链中复合物I的转录后调控,决定氧化磷酸化(OXPHOS)的速率。近些年,AIF被广泛研究。今天小编就带大家了解关于AIF调控氧化磷酸化支持肺癌的发展的文章AIF-regulated oxidative phosphorylation supports lung cancer development。

在本研究中,作者使用KrasG12D驱动的小鼠肺癌模型,通过AIF敲减、线粒体呼吸评估、糖酵解、WT或线粒体锚定的AIF重新表达和和肺癌转录组数据分析证明了AIF调控线粒体呼吸和OXPHOS,从而促进肺癌的进展。
结果:
1)减少AIF缺乏KrasG12D小鼠的肺癌
为了确定AIF在肺癌中的作用,我们将AIF fl/fl 小鼠与Lox-Stop-Lox-KrasG12D菌株杂交。Lox-Stop-Lox-KrasG12D小鼠在Cre缺失并诱导突变的KrasG12D等位基因后,逐步发展为非小细胞肺癌(NSCLCs),从上皮细胞增生到良性腺瘤和恶性腺癌。与对照组相比,KrasG12D驱动的肺癌模型中AIF的缺失显著延长了存活时间(图1a)。在所有时间点分析中,与对照相比,Aiffl / y KrasG12D小鼠肺中肿瘤区域的数量减少(图1 b, c)。这些结果表明,AIF基因失活可显著降低krasg12d驱动的肺癌发生。接下来我们评估了肿瘤的发生和肺癌的恶性进展。Ad5-CMV-Cre吸入4周后,微CT显示Aiffl/y KrasG12D小鼠肿瘤病灶明显减少,且在整个观察期间均观察到肿瘤病灶减少(图1d)。

2)AIF的缺失会损害OXPHOS并破坏线粒体结构
由于AIF影响肿瘤的发生和早期发展,我们从Aiffl/y KrasG12D和AIF+/y KrasG12D胎鼠中分离出原代肺细胞,随后用Ad5-CMV-Cre-eGFP感染培养细胞,激活KrasG12D表达,同时删除AIF(图2a)。正如之前研究中所预期的那样,AIF的缺失也导致线粒体复合物I蛋白的减少(图2a)。然后分析AIF突变原代肺细胞的耗氧量(OCR),以评估线粒体呼吸。Aiffl/y KrasG12D细胞在较低的基础水平上消耗氧气,产生更少的ATP,并且在对FCCP的响应中表现出有限的OCR增加,导致最大呼吸容量降低(图2 b,c)。
在体内Ad5-CMV-Cre感染6周后,我们从Aiffl/y KrasG12D和Aif+/y KrasG12D胎鼠中分离出原代转化的肺细胞。值得注意的是,小鼠感染的病毒滴度是正常浓度的5倍,以确保大多数肺细胞被感染(图2d)。与我们的短期培养相似,体内Ad5- CMV-Cre处理导致基础呼吸、ATP生成、最大呼吸和备用呼吸能力减少(图2e)。我们还对Ad5-CMV-Cre吸入16周后Aiffl/y KrasG12D和Aif+/y KrasG12D小鼠新分离肿瘤组织中的线粒体进行了表征。具有AIF活性的肿瘤细胞与缺乏AIF的肿瘤细胞线粒体数量无明显差异;然而,Aiffl/y KrasG12D肿瘤细胞线粒体肿胀,嵴结构明显减少,排列紊乱(图2f)。因此,AIF的缺失不仅会导致OXPHOS的普遍受损,还会导致线粒体形态异常。
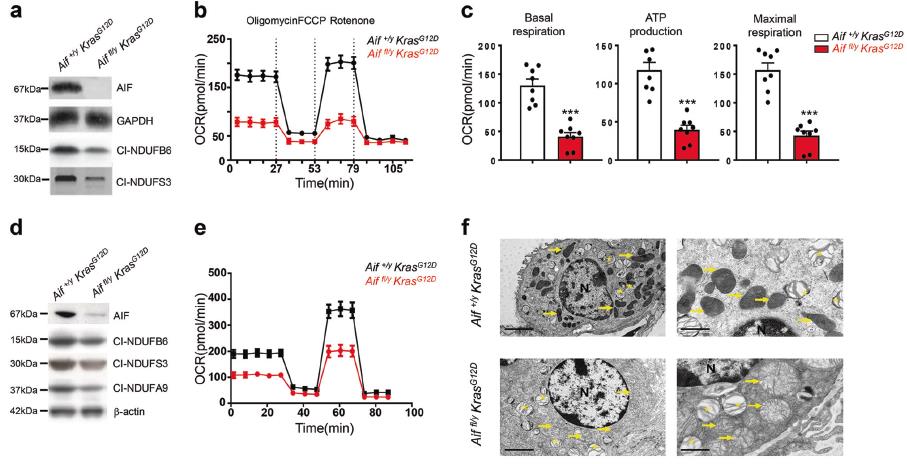
3)AIF缺乏增强糖酵解和对葡萄糖缺乏的敏感性
在添加葡萄糖刺激糖酵解后,Aiffl/y KrasG12D肺细胞的基础细胞外酸化率高于Aif+/y KrasG12D对照细胞(图3a)。添加寡霉素后,通过OXPHOS抑制ATP产生,Aiffl/y KrasG12D细胞和Aif+/y KrasG12D细胞的糖酵解活性均增加,但Aif+/y KrasG12D细胞的糖酵解活性明显增加(图3 b)。在体外Ad5-mSPC-Cre处理的Aiffl/y KrasG12D肺细胞中,糖酵解也增加了(图3 c,d)。对照Aif+/y KrasG12D细胞不受2-DG 72 h孵育的影响,,而Aiffl/y KrasG12D细胞死亡率增加(图3e)。同样,葡萄糖缺乏对Aif+/y KrasG12D细胞无明显影响,但影响Aiffl/y KrasG12D细胞的生长(图3f)。最后,我们观察到在葡萄糖缺乏条件下培养的Aiffl/yKrasG12D肺细胞内ATP水平显著下降(图3g)。这些结果表明AIF的缺失导致OXPHOS缺乏症和细胞代谢向糖酵解的转变。Aiffl/y KrasG12D肺细胞中,OXPHOS对ATP合成的贡献较小,主要由糖酵解提供,而Aif+/y KrasG12D细胞中的ATP大部分由OXPHOS生成。
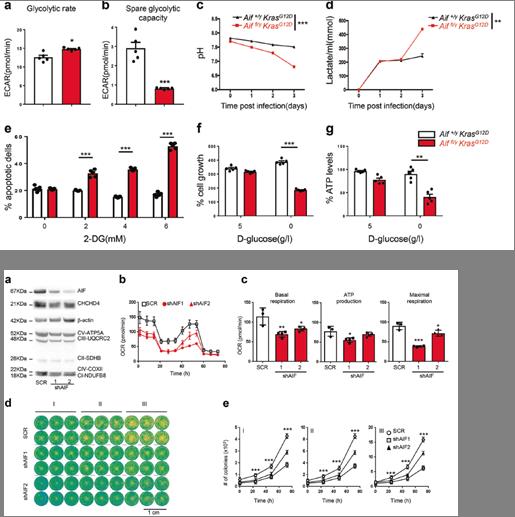
4)稳定的AIF缺失会损害KRAS WT和KRAS突变的肺癌细胞的克隆潜能和增殖
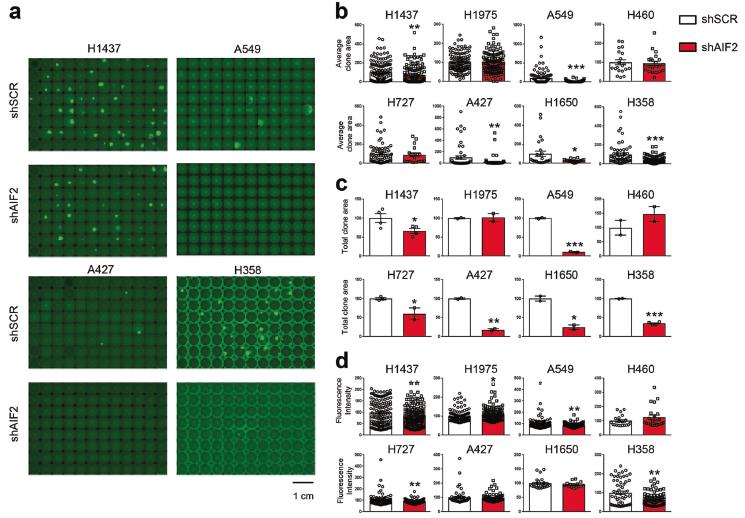
在KRAS突变的NSCLC A549细胞中,AIF的稳定下调抑制了CHCHD4的表达,并导致呼吸链蛋白如CIII-UQCRC2、cii - coxii和CI-NDUFB8的减少以及OXPHOS、ATP生成、克隆潜能和增殖的减少。另外,AIF敲除显著降低了H1437、H727、A427、H1650、H358以及A549细胞的克隆能力(图5)。
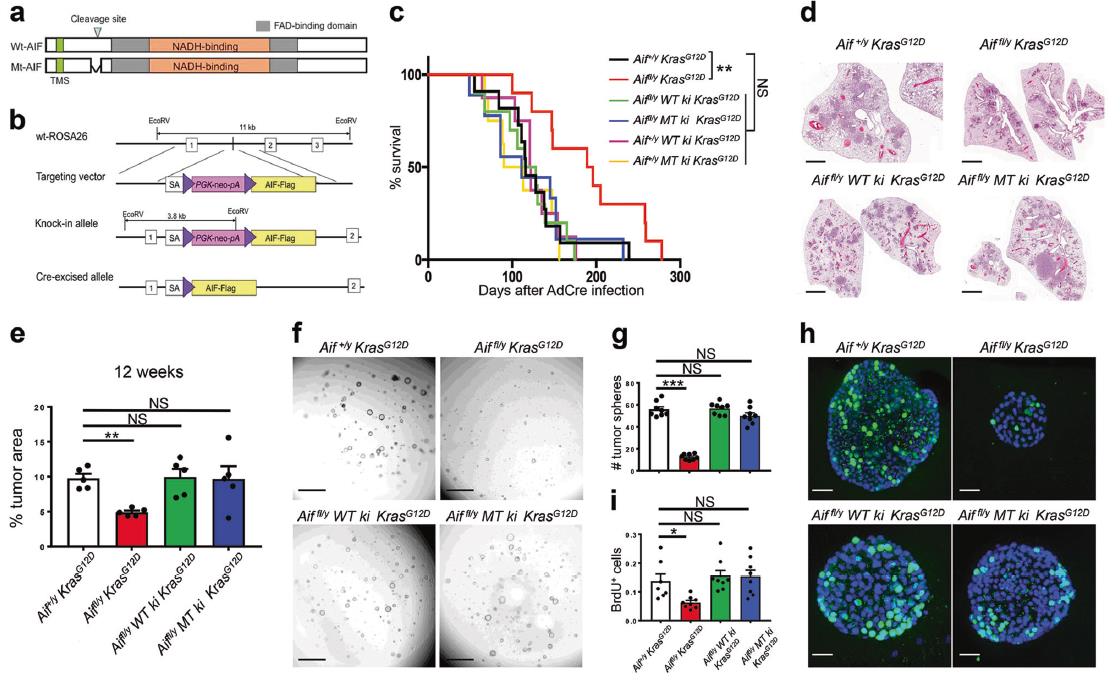
5)重新表达WT或线粒体锚定的AIF可恢复肺癌敏感性
Aiffl/y KrasG12D小鼠在吸入Ad5-CMV-Cre后存活的时间明显长于对照组小鼠。在敲除AIF的小鼠中重新表达WT AIF后,存活率显著降低。而且线粒体锚定突变体AIF替代内源性AIF也恢复了Aiffl/y KrasG12D小鼠的癌症表型(图6c)。另外WT或突变AIF的重新表达导致肺肿瘤负荷增加(图6d,e)。接下来为了进一步研究AIF和OXPHOS在肿瘤细胞增殖和肿瘤干细胞样特性中的功能相关性,我们采用最近开发的3D肿瘤球体培养法,从Ad5-mSPC-Cre感染6周后的Aif+/y KrasG12D、Aiffl/y KrasG12D、Aiffl/y WT ki KrasG12D、Aiffl/y MT ki KrasG12D小鼠中分离纯化的原发肺细胞,进行细胞培养。在从每个基因型中植入相同数量的肿瘤细胞后,我们发现Aiffl/y KrasG12D小鼠与Aif+/y KrasG12D小鼠相比,来自Aiffl/y KrasG12D小鼠的肿瘤球体数量显著减少。WT和突变型AIF的重新表达几乎完全恢复了茎样性质,形成肿瘤球形,达到AIF +/y KrasG12D对照水平(图6f,g)。接下来,我们通过BrdU标记来确定这些肿瘤球的增殖能力。再次,我们观察到Aiffl/y KrasG12D肿瘤球体中阳性细胞明显少于AIF WT、AIF WT敲入和AIF突变敲入肿瘤球体(图6h,i)。
6)WT或线粒体锚定AIF的修复可消除线粒体呼吸缺陷
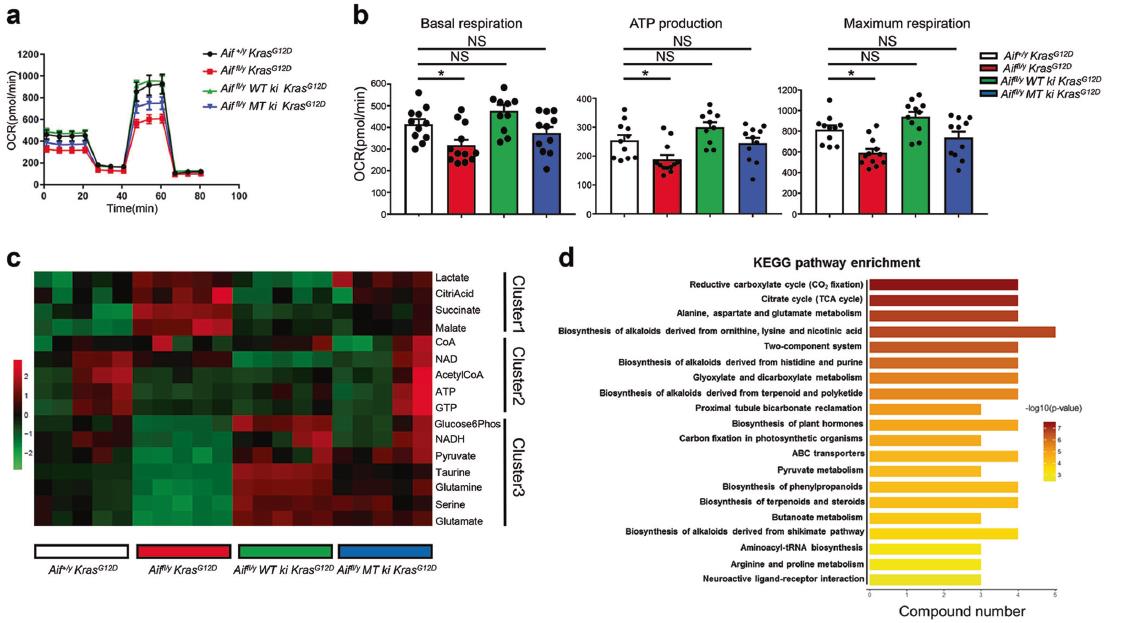
将WT或线粒体锚定的AIF重新引入Aiffl/y KrasG12D肺细胞中,可将其呼吸能力提高到与AIF +/y KrasG12D对照组相当的水平(图7a, b)。另外,与生物能量谱一致,AIF缺陷细胞所含PEP和NADH明显减少,但乳酸含量高得多(图7c),这表明AIF耗尽后OXPHOS确实受到抑制。另外,三羧酸循环的中间代谢物,包括柠檬酸、苹果酸和琥珀酸,在AIF缺乏的细胞中大量增加(图7c),这与KEGG富集分析一致(图7d)。
7)AIF在人类肺肿瘤中经常过度表达,高AIF表达与低生存率有关
最后,我们通过分析已发表的数据集,探讨AIF在人类肺癌中的临床意义。肺癌和不同肺癌亚型的转录组研究表明,与正常组织相比,肺癌组织中AIF mRNA水平显著上调(图8a)。为了在蛋白水平上验证这一发现,我们用免疫组织化学方法研究了AIF在人类肺癌样本中的表达。与之前的研究结果一致,我们观察到Kras突变和Kras WT肺癌患者的肿瘤组织中AIF表达均高于正常肺组织(图8b)。另外,我们发现,在三个不同的NSCLC群体中,不仅AIF,而且编码复合物I亚基或其组装因子的绝大多数基因在NSCLC组织中都比正常相邻肺组织明显过表达(图8c)。因此,当我们用Ad5-mSPC-Cre感染Aif+/y和Aif+/y KrasG12D小鼠的肺细胞时,在激活KrasG12D 8天后检测到AIF表达显著诱导;然而,在表达WT Kras的非转化性肺细胞中没有观察到这种效应(图8d)。此外,我们通过分析TCGA数据集中的RNAseq数据,评估了AIF表达与总生存率之间的相关性。发现无论KRAS突变情况如何,AIF高表达与预后不良呈负相关(图8e)。同样,免疫组化检测显示,NSCLCs表达低水平AIF蛋白的患者比AIF高肿瘤患者生存时间更长(图8f),这与AIF高表达与预后不良相关的结论一致。