






PUMA通过减少FASN泛素化促进人类透明细胞肾细胞癌的脂质积累和肿瘤进展
p53上调凋亡调节因子(PUMA)传统上被认为可在多种癌症中促进细胞凋亡并增强化疗疗效,但由于透明细胞肾细胞癌(ccRCC)对化疗具有耐药性,其在ccRCC中的作用尚不明确。该研究通过分析公共数据集、临床组织样本和细胞系实验,发现PUMA在ccRCC中具有新型致癌作用,这与其已知的凋亡功能不同。PUMA的异常高表达与临床分期呈正相关,且预示预后不良。值得注意的是,PUMA在ccRCC中的作用似乎独立于凋亡过程,相反,它通过涉及关键代谢调节因子 —— 脂肪酸合成酶(FASN)的机制促进肿瘤进展和脂质积累。具体而言,PUMA的N44-102 氨基酸序列(不同于先前研究的BH3结构域)是其与FASN相互作用的关键。机制上,PUMA通过与泛素特异性蛋白酶 15(USP15)结合来稳定 FASN,减少FASN的泛素化和降解,从而形成PUMA-USP15-FASN轴。这些发现挑战了PUMA在癌症生物学中的既定认知。此外,敲低PUMA可显著抑制肿瘤生长,并增强ccRCC肿瘤对代谢抑制的敏感性。这些结果表明,PUMA是一种新型代谢调节因子,可能成为ccRCC的治疗靶点。联合抑制PUMA和FASN进一步支持了靶向这一代谢轴的治疗潜力。这篇文章于2025年6月发表在《cell death & disease》期刊上,IF:9.6。
研究技术路线:
主要实验结果:
1、PUMA 在 ccRCC 中的高表达与临床分期呈正相关
作者从 GEPIA 和 UCSC Xena 数据中心获取了 PUMA 的 mRNA 数据。GEPIA 的基因点图分析显示,与正常组织相比,PUMA 在 11 种癌症类型中均呈高表达(见图 1A),包括 ccRCC(即肾透明细胞癌,KIRC)。基于 534 例 ccRCC 患者和 72 对配对组织样本(肿瘤及相应的正常肾组织)的数据,小提琴图和折线图也显示 ccRCC 肿瘤中 PUMA mRNA 水平较高(图 1B、C)。在 TCGA 数据集中,将 534 例 ccRCC 样本中 RNA 水平高于中位数(7.5085)的定义为高表达,低于中位数的定义为低表达(图 1D)。部分 Kaplan-Meier 曲线分析表明,PUMA 高表达可能与总生存期缩短相关(图 1D 和 S1A)。通过比较 ccRCC 肿瘤组织与正常肾组织的曲线下面积(AUC)值,发现 PUMA 的表达水平具有高特异性和临床诊断价值(图 1E)。小提琴图证实,PUMA 高表达与 ccRCC 的晚期肿瘤分期和癌症分级相关(图 1F-H)。单因素 Cox 分析显示,PUMA mRNA 水平与多种临床病理参数显著相关,包括 TNM 分期。然而,多因素 Cox 分析表明,PUMA 并非独立的预后标志物(p=0.194)。
图 1:PUMA 在 ccRCC 中的高表达与临床分期呈正相关。
为进一步验证作者的发现,作者检查了 25 对 ccRCC 患者的临床组织样本。实时定量 PCR(qPCR)分析结果显示,肿瘤组织中 PUMA mRNA 表达较正常组织显著增加(图 1I)。免疫组织化学分析再次证实了肿瘤组织中 PUMA 的高表达(图 1J)。此外,Western blot 分析显示,在 22 对组织样本中,肿瘤组织的 PUMA 蛋白表达高于正常组织(图 1K,L)。另外,与人类肾细胞(HK-2)相比,人类 ccRCC 细胞(A-498、ACHN、Caki-1)的 PUMA 蛋白和 mRNA 表达均较高(图 1M,N)。值得注意的是,786-O 细胞的 PUMA mRNA 表达显著高于 HK-2,而 786-O 与 OS-RC-2 之间的蛋白水平无显著差异(p>0.05)。A-498 作为典型的 ccRCC 细胞系,Caki-1 代表转移性 ccRCC。这两种细胞系在 VHL 和 p53 上表现出不同的突变模式。作者选择它们在后续实验中进行比较,以强调结果的普遍性。总之,基于公共数据、临床组织和细胞系的所有评估均证实,PUMA 在 ccRCC 中高表达,且与患者的临床分期较高相关。
2、PUMA 在 ccRCC 中的新型致癌作用与凋亡无关
为了研究 ccRCC 中高表达 PUMA 的生物学作用,作者将携带 PUMA 特异性短发夹 RNA(shPUMA)的慢病毒质粒转染到 A-498 和 Caki-1 细胞中,以建立稳定的 PUMA 敲低细胞系。通过 Western blotting 和 qPCR 验证,敲低效率超过 70%。对照组(CN)使用空白慢病毒质粒生成。测量关键凋亡通路分子以评估 PUMA 表达与凋亡的相关性,包括全长 PARP、Bcl-2、caspases-3、-6 和 - 7,以及线粒体凋亡标志物如细胞色素 C。Western blot 结果显示,在对照组和 shPUMA 组之间,无论是全细胞裂解物还是细胞质 / 线粒体组分,凋亡标志物的表达或激活均无显著差异。此外,细胞色素 C 保留在线粒体中,进一步支持没有线粒体凋亡激活。对多种凋亡标志物蛋白的比较证实,在作者的模型中,PUMA 水平与凋亡活性没有直接相关性。
为了研究 PUMA 是否影响 ccRCC 的进展,作者对 PUMA 敲低组和对照组进行了集落形成实验和细胞活力测试。结果表明,与对照组相比,PUMA 敲低组在 A-498 和 Caki-1 细胞系中的细胞增殖率和集落形成能力显著降低(图 2A-C)。Transwell 实验显示,与对照组相比,PUMA 敲低组的细胞迁移和侵袭能力减弱(图 2D,E)。总之,PUMA 的敲低与 A-498 和 Caki-1 细胞系恶性进展的显著降低相关。
图 2:PUMA 在 ccRCC 中的新型致癌作用与细胞凋亡无关。
ccRCC 的恶性进展由代谢重编程驱动,其特征是细胞质中糖原和脂滴的大量积累。如图 S1F 所示,PUMA 与 ccRCC 中的葡萄糖积累没有直接关联,但作者观察到其与脂滴呈正相关。与对照组相比,PUMA 敲低组在 A-498 和 Caki-1 细胞系中的脂滴含量均降低。结果通过油红 O(ORO)染色和定量分析呈现(图 2F,G)。与对照组相比,PUMA 敲低组中 A-498 和 Caki-1 细胞的甘油三酯和总胆固醇均降低(图 2H),这在 ccRCC 的背景下呈现出独特的模式。相反,在 PUMA 过表达组中,细胞增殖、脂滴、甘油三酯和总胆固醇均增加。这些发现暗示 PUMA 在促进 ccRCC 进展和脂质积累中起作用。
为了证实作者之前的体外发现,作者在裸鼠中创建了皮下和转移性肿瘤模型。在 PUMA 敲低组和对照组中均进行了皮下和尾静脉注射。大体肿瘤图像和生长曲线显示,与对照组相比,PUMA 敲低组的肿瘤大小、重量和生长速率均降低(图 2I-K)。免疫组织化学显示,敲低组的 PUMA 表达显著降低,各组之间的凋亡标志物无差异。H&E 和 ORO 染色一致显示,PUMA 敲低组的组织切片恶性程度较低,脂质积累低于对照组(图 2L)。在肝脏大体图中,与对照组相比,PUMA 敲低组的肝转移灶更少(图 2M)。总的来说,作者进一步巩固了之前的发现(图 2A-H),即 PUMA 促进 ccRCC 中的肿瘤进展和脂质积累,且独立于其在凋亡中的作用。
近期研究强调线粒体凋亡通路的激活是肿瘤生长和转移的驱动因素。这提出了 PUMA 可能通过亚致死信号促进 ccRCC 侵袭性肿瘤行为的可能性。按照之前的方法,作者评估了线粒体和核组分中的 EndoG 水平。与对照组相比,shPUMA 组中未观察到 EndoG 的显著核转位或激活。同样,通过 ICAD(CAD 的抑制剂)和 γH2AX(DNA 损伤标志物)分析 CAD 活性,显示 shPUMA 组和对照组之间无显著差异。这些结果表明,通过 EndoG 或 CAD 的亚致死信号不参与观察到的肿瘤表型。然后,作者使用 CRISPR-Cas9 生成 ENDOG/CAD 和 BAK/BAX 双敲除(DKO)细胞,通过 Western blotting 验证敲除效率。对这些 DKO 细胞中肿瘤细胞生长的评估显示,shPUMA 组和对照组之间无表型差异。因此,与 PUMA 相关的侵袭性生长表型独立于通过 EndoG/CAD 的亚致死信号或其通过 BAX/BAK 的促凋亡功能,表明 PUMA 通过其他机制发挥作用。
3、直接相互作用:FASN 是 PUMA 功能的关键
为了探索 PUMA 在 ccRCC 中的亚细胞定位,作者使用细胞质 - 核分级分离和线粒体分离实验分析了其在不同细胞区室中的表达。Western blotting 显示,PUMA 主要定位于 HK-2、A-498、Caki-1 和 786-O 细胞系的线粒体中。免疫荧光成像证实了细胞质 PUMA 的定位及其与线粒体标志物的共定位,与之前的研究一致。这些发现表明,鉴于线粒体在肿瘤代谢重编程(尤其是脂质代谢)中的关键作用,PUMA 可能参与 ccRCC 的脂质代谢。
为了进一步探索 PUMA 在 ccRCC 内的分子相互作用,作者使用 PUMA 抗体进行免疫沉淀(IP)实验,从全细胞和细胞质裂解物中分离蛋白质。全细胞蛋白分析显示,与 IgG 抗体相比,PUMA 抗体下拉了 320 种蛋白质,表明 PUMA 与不同的蛋白质池相互作用。基于 IP 和 MS 结果,作者进行了基因本体论(GO)功能富集和京都基因与基因组百科全书(KEGG)通路分析。结果表明,PUMA 相关分子在代谢通路和脂质代谢通路中富集。为了提高检测灵敏度,进行了细胞质分级分离以从细胞质中分离蛋白质。用 PUMA 抗体进行免疫沉淀从细胞质组分中获得了 222 种蛋白质,而用 IgG 下拉了 139 种蛋白质。通过将全细胞 IP、细胞质 IP 及其各自的 IgG 对照的数据集相交,确定了 43 个潜在靶点。将细胞质 IP 数据集与脂质代谢数据进一步相交,作者发现了 28 个与脂质代谢相关的分子。值得注意的是,这 28 个分子是全细胞和细胞质数据集中 43 个靶点的一部分,验证了它们在脂质代谢通路中的相关性。在作者的分析中,FASN 在与脂质代谢通路相关的分子中综合得分最高。鉴于线粒体为脂肪酸合成提供乙酰辅酶 A 和柠檬酸,而 FASN 利用乙酰辅酶 A 作为其脂肪酸生物合成的底物,因此提出 FASN 是参与脂质代谢的 PUMA 的关键下游靶点是合理的。
作者进一步研究了 ccRCC 中与脂质代谢相关的 PUMA 分子相互作用。基于 IP 和 MS 结果,作者重叠了三个数据集:细胞质 IgG、细胞质 IP 和脂肪酸生物合成。该分析确定 FASN(脂肪酸生物合成中的关键酶)是仅有的与 PUMA 相互作用的分子(图 3A)。这一发现与图 S4G 一致,表明 FASN 是 PUMA 在脂质代谢中作用的关键。内源性和外源性 Co-IP 实验证实了 A-498、Caki-1 和 HEK293T 中 PUMA-FASN 的相互作用(图 3B,C)。此外,BAX/BAK DKO 细胞中的 Co-IP 实验进一步验证了这种相互作用,表明 PUMA 与 FASN 的结合独立于其促凋亡功能。共聚焦成像证实 FASN 主要定位于细胞质(图 3D),并与 PUMA 共定位,尤其是在线粒体区域。这些结果突显了 FASN 在 PUMA 介导的 ccRCC 脂质代谢中的重要作用。
图 3:直接相互作用:FASN 是 PUMA 功能的关键。
接下来,作者使用 FRET 分析和 Co-IP 测定验证了 PUMA-FASN 相互作用。作者构建了 Clover2-PUMA 和 mRuby2-FASN 质粒,通过分别用一对 FRET 荧光团 Clover(供体)和 mRuby2(受体)标记 PUMA 和 FASN。FRET 测定结果证实了 PUMA 和 FASN 之间的直接相互作用(图 3E,F)。作者生成了一系列 PUMA 缺失构建体(HA-PUMA ΔBH3、HA-PUMA ΔMLS、HA-PUMA ΔBH3/MLS、HA-PUMA-N43、HA-PUMA-N44–102、HA-PUMA-N103–140),并将它们与 FLAG-FASN 质粒共转染到 HEK293T 细胞中(图 3G)。Co-IP 结果显示,PUMA 的 N44-102 和 N103-140 区域对其与 FASN 的相互作用至关重要(图 3H,I)。这不同于先前公认的 BH3 结构域,后者被认为是主要的相互作用基序。最近的一篇论文提出了另一种观点,认为 PUMA 的结合结构域可能超出传统的 BH3 基序,其亚细胞定位可能受其表达水平和结合结构域的影响。这与作者的发现一致,即 PUMA 在其 BH3 结构域之外与 FASN 相互作用,表明参与 PUMA 在脂质代谢中作用的替代结合位点。总之,作者发现 ccRCC 中 PUMA 和 FASN 之间存在蛋白质 - 蛋白质相互作用,PUMA 的 N 端 44-140 序列在这种相互作用中起关键作用。
4、PUMA 介导 FASN 表达以驱动 ccRCC 进展并促进脂质积累
FASN 在脂肪酸生物合成中至关重要,并与包括 ccRCC 在内的各种癌症的肿瘤发生和进展相关。作者进行了 Transwell 实验以探索 PUMA 和 FASN 之间的调节关系,发现 FASN 的补充几乎恢复了 PUMA 敲低组的迁移和侵袭能力(图 4A,B)。在细胞活力测定中观察到类似结果(图 4C)。这些发现表明,PUMA 调节 FASN 的致癌活性,从而促进 ccRCC 中的肿瘤增殖和迁移。同样,ORO 染色和甘油三酯及胆固醇的测量显示,FASN 的补充几乎恢复了 PUMA 敲低组的脂滴形成和脂质代谢物水平(图 4D-F)。这些结果与 Transwell 实验结果一致(图 4A,B),进一步支持 PUMA 通过 FASN 促进 ccRCC 中脂质代谢和恶性进展的作用。
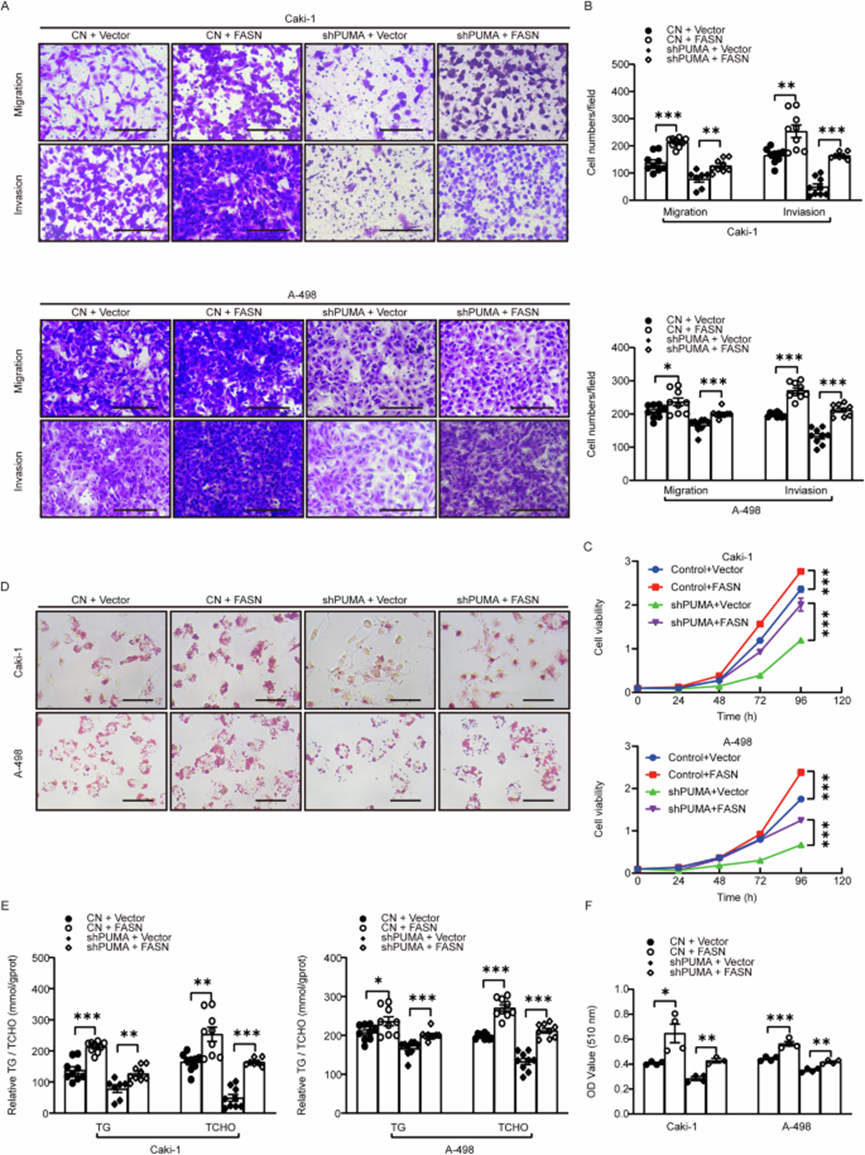
图 4:PUMA 介导 FASN 表达以驱动 ccRCC 进展并促进脂质积累。
为了进一步研究 PUMA-FASN 相互作用的功能相关性,作者在 PUMA 敲低细胞中过表达全长 PUMA 和截短构建体(N44-102 和 N103-140)。Transwell 实验显示,PUMA 的补充恢复了细胞迁移,N44-102 构建体产生了类似的效果。细胞增殖和 ORO 染色测定再次证实,全长 PUMA 恢复了增殖能力和脂滴水平,添加 N44-102 基序达到了类似的结果。这些结果进一步强调了 PUMA 的 N44-102 基序在与 FASN 直接相互作用中的关键作用,从而促进癌细胞增殖和脂质积累。
5、PUMA 通过泛素 - 蛋白酶体途径增强 FASN 稳定性和表达
作者进一步探索揭示 PUMA 和 FASN 之间相互作用的机制。基于不同临床分期(I-II 和 III-IV)的 10 对临床组织样本进行免疫组织化学染色。研究结果显示,随着恶性程度的升高,PUMA 和 FASN 的染色均增强(图 5A)。重要的是,观察到 PUMA 和 FASN 染色强度之间存在正相关(见图 5A)。随后,在 12 对 ccRCC 临床组织样本中检查了蛋白质表达。观察到 FASN 和 PUMA 的蛋白质表达之间存在正相关,与正常组相比,PUMA 敲低组中 FASN 表达显著降低(图 5B,C)。在体内,FASN 表达随着 PUMA 敲低而相应降低(图 5D)。将研究扩展到 ccRCC 细胞系,Western blotting 显示 FASN 和 PUMA 的蛋白质水平协同升高(图 5E)。FASN 的蛋白质和 mRNA 表达水平在 ccRCC 细胞系中高度升高,类似于 PUMA。随后的 Western blot 分析表明,与对照组相比,PUMA 敲低组中 FASN 蛋白表达降低,而过表达 PUMA 细胞系中 FASN 蛋白表达增加(图 5F,G 和 S6C,D)。在 qPCR 分析中,无论 PUMA 表达如何,FASN mRNA 水平均未观察到显著变化(图 5G 和 S6D)。这些发现与之前的研究一致,表明 FASN 表达主要在翻译后水平而不是 mRNA 水平受到调节。最后,与对照组相比,蛋白质稳定性测定显示 PUMA 敲低组中 FASN 的半衰期较短(图 5H,I)。
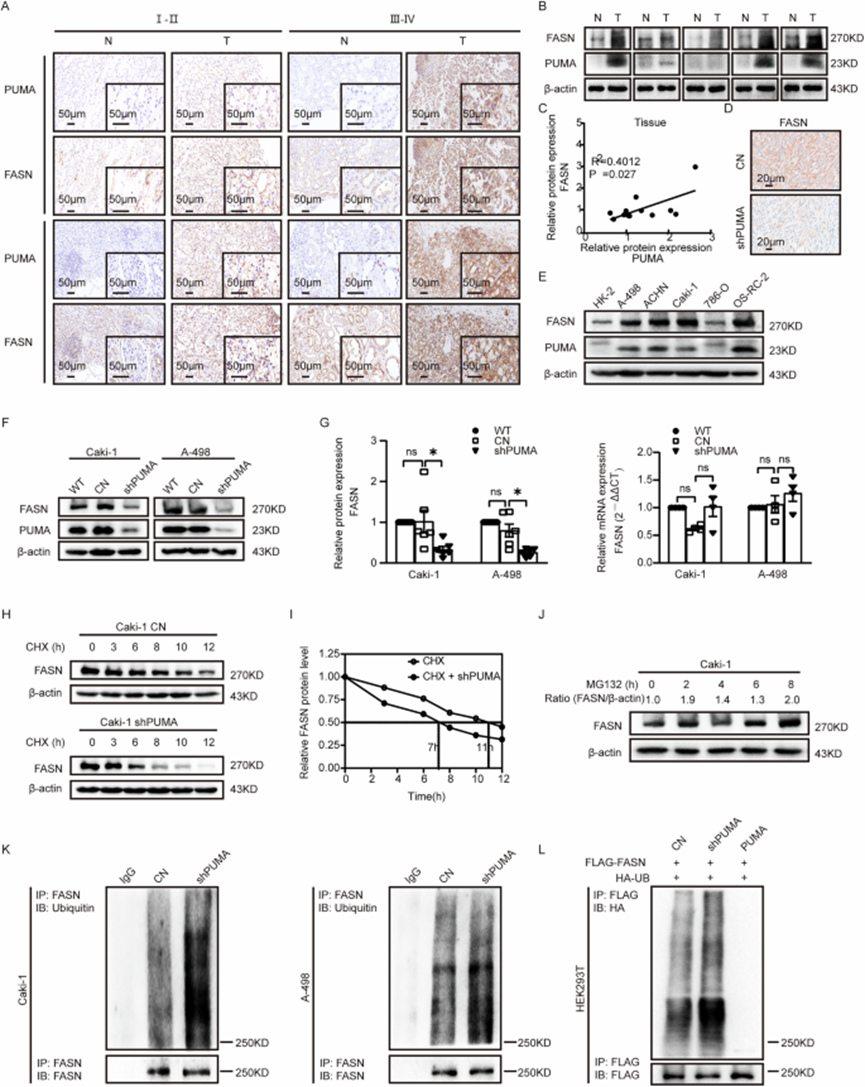
图 5:PUMA 通过泛素-蛋白酶体途径增强 FASN 稳定性和表达。
最近的研究报道了细胞凋亡诱导与脂质代谢之间的相关性,表明 FASN 过表达也可能与细胞凋亡信号传导相关。为了研究 PUMA 是否独立于其促凋亡活性调节 FASN,作者评估了 BAX/BAK DKO 细胞中对照组和 PUMA 过表达组的 FASN 蛋白水平。Western blot 分析显示,PUMA 过表达仍然导致 BAX/BAK DKO 细胞中 FASN 表达增加,与 BAX 和 BAK 完整的细胞中的观察结果一致。这些发现证实,PUMA 调节 FASN 独立于其促凋亡功能。
为了研究参与 FASN 蛋白降解的途径,作者用两种经典的蛋白质降解途径抑制剂处理 ccRCC 细胞系:MG132(泛素 - 蛋白酶体抑制剂,10μM)和氯喹(自噬 - 溶酶体抑制剂,25μM)。时间梯度 Western blotting 显示,MG132 处理增加了 FASN 蛋白水平(图 5J 和 S6F,G)。PUMA 敲低组和对照组用cycloheximide(CHX,蛋白质合成抑制剂,25μM)处理。12 小时后,每组中的一半细胞用 MG132 处理,其余一半不接受额外处理。如图 S6H 所示,添加 MG132 导致两组中 FASN 蛋白表达水平增加。为了进一步研究 PUMA 如何调节 FASN,作者进行了泛素化 IP 测定,结果显示与对照组相比,PUMA 敲低组中 FASN 泛素化增加(图 5K,L)。作者建立了稳定敲低 PUMA 的 HEK293T 细胞系,通过 Western blotting 证实。在 HEK293T 细胞中,PUMA 过表达降低了 FASN 泛素化水平(图 5L)。这些结果表明,PUMA 通过泛素 - 蛋白酶体途径调节 FASN 的泛素化并调控其蛋白质稳定性。
6、PUMA-USP15-FASN 轴及其与 FASN 抑制剂的协同效应
由于 PUMA 缺乏催化活性结构域,可能需要一种中间蛋白来促进其对 FASN 泛素化的调节。回顾之前的 MS 分析结果,作者发现去泛素酶 USP7 和 USP15 可能与 PUMA 相互作用(图 6A)。然而,关于 EIF3H 作为去泛素化酶的理解和应用仍不完整,需要进一步全面探索。在 HEK293T 细胞中,作者进行了以下组合的共转染:(1)FLAG-PUMA 和 HA-USP7 质粒,以及 FLAG-PUMA 和 HA-USP15 质粒;(2)FLAG-FASN 和 HA-USP7 质粒,以及 FLAG-FASN 和 HA-USP15 质粒。Co-IP 结果表明,USP15 是 PUMA 和 FASN 之间的调节衔接蛋白(图 6B)。
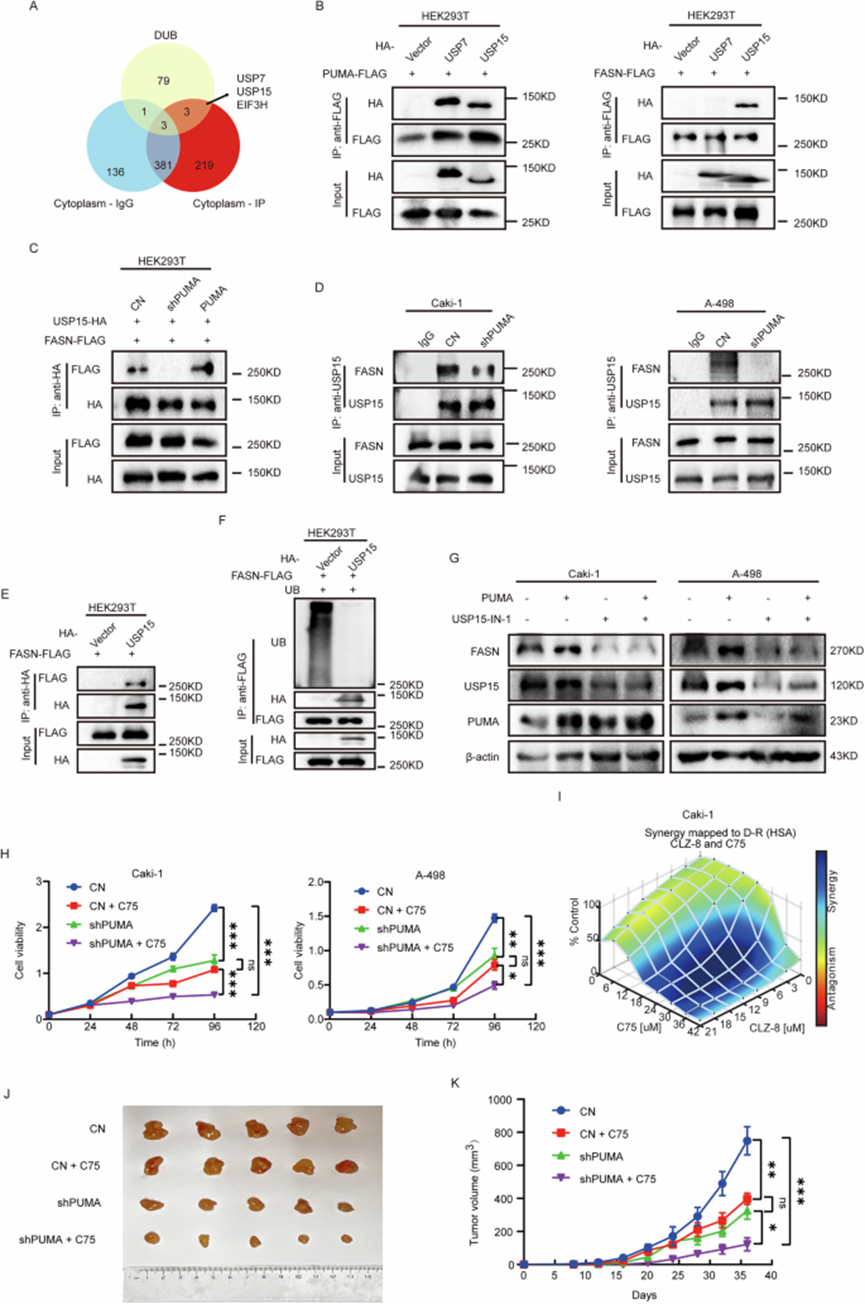
图 6:PUMA-USP15-FASN 轴及其与 FASN 抑制剂的协同作用。
通过免疫沉淀进一步验证了 PUMA-USP15-FASN 轴。在 PUMA 敲低细胞中,与对照组相比,与 USP15 共免疫沉淀的 FASN 量减少,而 PUMA 过表达增加了与 USP15 相关的 FASN 水平(图 6C)。在 Caki-1 和 A-498 细胞系中观察到类似结果,表明 PUMA 敲低组中 USP15 免疫纯化的 FASN 水平降低(图 6D)。此外,HEK293T 细胞中 USP15 过表达降低了 FASN 的泛素化水平,增强了其免疫纯化(图 6E,F)。USP15 抑制后,FASN 的蛋白质表达显著降低,与 PUMA 表达水平无关(图 6G)。综合这些结果,作者证实 PUMA 通过 USP15 调节 FASN 的泛素化,这巩固了 PUMA-USP15-FASN 轴存在的合理性。
先前的研究已将 PUMA 确立为抑制 ccRCC 恶性进展的有前景的治疗靶点。然而,靶向药物的不良反应和脱靶效应是当前 ccRCC 治疗中的难题。鉴于 FASN 抑制剂单独的临床疗效有限,越来越多的建议主张将其与其他靶点联合使用。因此,作者深入研究了 PUMA 和 FASN 抑制剂的联合疗效。作者评估了四个不同实验组中肿瘤细胞系的生长情况:对照组、PUMA 敲低组、用 C75(一种 FASN 抑制剂,35μM)处理的对照组,以及用 C75 处理的 PUMA 敲低组。PUMA 敲低组和用 C75 处理的对照组中肿瘤细胞增殖受到抑制,而同时接受 C75 和 PUMA 敲低的组中效果更显著(图 6H)。具体而言,PUMA-KD 组(与对照组相比)的抑制效率显著高于 PUMA 和 FASN 联合抑制组(与单独 FASN 抑制组相比)。使用 CLZ-8(一种 PUMA 抑制剂)和 C75 评估它们在 A-498 和 Caki-1 细胞中的联合效应。使用 Combenefit 软件分析两种抑制剂之间的相互作用。Combenefit 的分析显示 CLZ-8 和 C75 之间存在明显的协同效应(图 6I)。此外,剂量 - 反应分析表明,增加 CLZ-8 浓度允许减少 C75 剂量,同时保持疗效。这些发现强调了 PUMA 和 FASN 抑制的协同潜力,表明它们的联合治疗是临床中 ccRCC 的有前途的治疗方法。
因此,作者进行了额外的实验来验证敲低 PUMA 对裸鼠中抑制剂 C75 的增强作用。分组与体外实验一致(图 6H)。用 C75 处理的两组(包括图 6H 中的对照组和 PUMA 敲低组)每周腹腔注射 20mg/kg,而没有 C75 的对照组和 PUMA 敲低组腹腔注射等体积的生理盐水,共 36 天。结果与体外实验一致(图 6H)。与任何单一干预相比,PUMA 敲低组和 C75 联合使用显著减缓了肿瘤生长(图 6K)。这些发现表明,PUMA 在更广泛的调节网络中起作用,FASN 是关键但非仅有的组成部分。此外,PUMA 和 FASN 抑制的协同潜力支持它们的联合治疗作为临床中 ccRCC 的有前途的治疗策略。
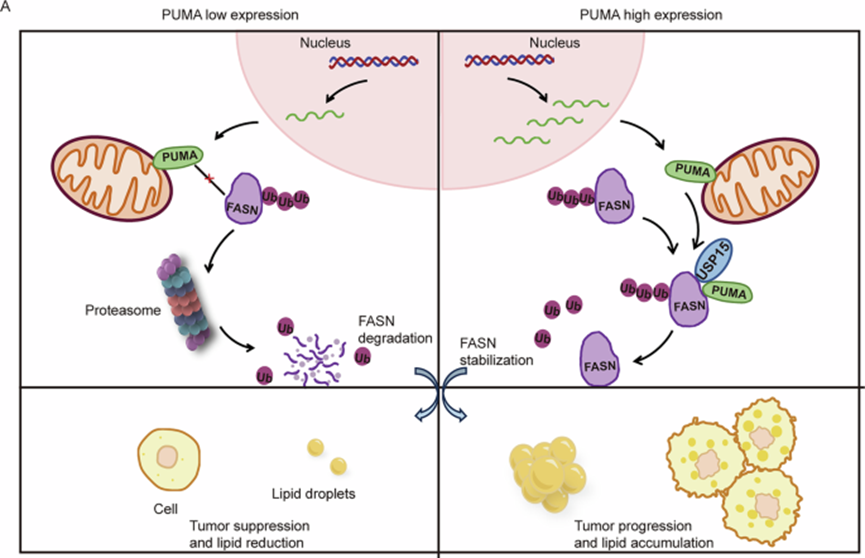
7、ccRCC 中 PUMA-USP15-FASN 轴的机制模型
总之,作者提出了 PUMA-USP15-FASN 轴的机制模型(图 7A)。该模型揭示,ccRCC 中 PUMA 的异常高表达调节 FASN,驱动细胞内脂质积累和肿瘤的恶性进展。作者的分子研究揭示了一种新的调节机制,其中 PUMA 通过 USP15 作为衔接蛋白与 FASN 结合,形成 PUMA-USP15-FASN 轴。该轴的建立有助于揭示 ccRCC 中的肿瘤进展和脂质代谢重编程。此外,与单独干预相比,PUMA-FASN 抑制的联合调节可以更有效地转化为实际临床应用。
参考文献
Luo, Q., Wang, Q., Shi, J. et al. PUMA reduces FASN ubiquitination to promote lipid accumulation and tumor progression in human clear cell renal cell carcinoma. Cell Death Dis 16, 460 (2025).
实验方法:
细胞实验:细胞培养及转染、Transwell 实验、细胞活力测定、集落形成实验
常规分子实验:WB、RT-qPCR、免疫组织化学与免疫荧光、油红 O(ORO)染色、PAS反应、甘油三酯和总胆固醇测定、免疫沉淀(IP)与质谱(MS)分析、共免疫沉淀(Co-IP)实验、荧光共振能量转移(FRET)分析
动物模型及病理分析:体内异种移植模型