






人18S rRNA m6A甲基转移酶新发现——METTL5
RNA甲基化在mRNA上被大量发现,rRNA甲基化修饰却研究的很少,并且rRNA甲基化修饰作用仍不清楚。近日,来自法国巴黎理工学院的研究团队在Nucleic Acids Research期刊上发表题名为:The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112的研究文章。该文章主要发现METTL5一个是负责18S rRNA m6A修饰的酶,并确认ZCCHC4为28S rRNAm6A修饰酶;METTL5必须与已知的甲基转移酶激活剂TRMT112形成异源二聚体复合物,以在细胞中获得代谢稳定性。文章还提供了METTL5-TRMT112的第一个原子分辨率结构。

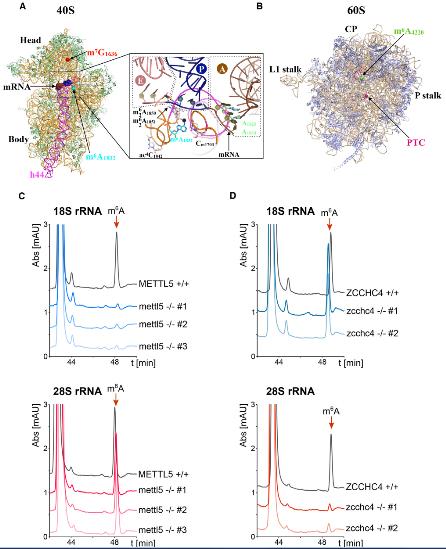
1.鉴定m6A1832和m6A4220上沉积的18S和28S rRNA甲基转移酶
为了鉴定在18S rRNA的A 1832和28S rRNA的A 4220处沉积m 6 A 的甲基转移酶,在UniProt数据库中搜索了具有N6-腺苷特异性DNA甲基转移酶。检索到七个结果结果:HEMK1、HEMK2、TRMT11、MGAM2、METTL4、METTL5和ZCCHC4。根据人类蛋白质图谱计划,METTL4是人类U2-OS细胞中线粒体蛋白;METTL5和ZCCHC4分别位于核仁和细胞核,即核糖体生物发生的初始步骤。有研究报道,人类METTL5与RNA特异性结合,但不与DNA结合。这些结果促使作者研究了METTL5和ZCCHC4是否可能是导致人类rRNA上m 6 A沉积的酶。
为了验证这一假设,通过CRISPR-Cas9基因编辑消除了人类细胞(HCT116)二倍体两个等位基因上蛋白质的部分编码序列,从METTL5中删除了外显子3,从ZCCHC4中删除了外显子7。三个独立的mettl5-/-细胞系和两个独立的zcchc4-/-细胞系,通过PCR验证基因组DNA上两个等位基因的缺失。为了测试METTL5和ZCCH4在核糖体RNA修饰中的作用,通过离心从不同的mettl5-和zcchc4缺失的细胞系以及对照HCT116细胞中纯化了成熟的18S和28S rRNA,然后将其消化成核苷并用HPLC定量分析。在对照细胞中,m6A核苷很容易在纯化的18s和28s中检测到,48分钟洗脱。在所有三个mettl5-/-细胞中,18S rRNA m6A峰丢失,而28S rRNA m6A峰保持不受影响。相反,在两个zcchc4-/-克隆中,18S rRNA m6A峰不变,而28S rRNA m6A峰消失。这些数据可以得出结论, METTL5和ZCCHC4是人类甲基转移酶,分别负责在18S rRNA上形成m 6 A 1832和在28S rRNA 上形成m 6 A 4220。ZCCHC4鉴定为28S rRNA m6A修饰酶与最近的文献报道完全一致。
2.METTL5和ZCCHC4可作用于细胞生长和核糖体生物发生
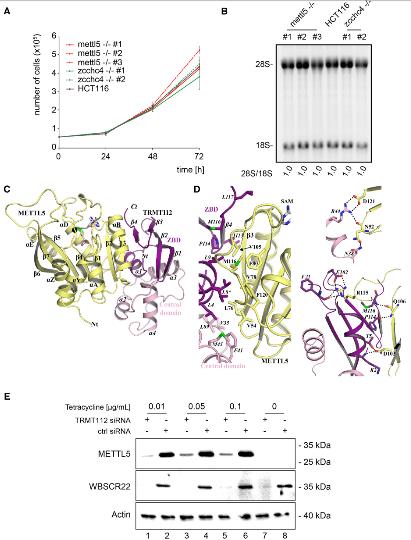
确定了18S和28S rRNA m 6 A甲基化酶后,检查这些修饰是否对细胞生长和核糖体生物发生有影响。首先,利用活体细胞染色计数监测细胞增殖,实验结果表明无METTL5或ZCCHC4基因的对细胞生长没有显著影响。接下来,从HCT116细胞和ETTL5-/-和ZCCHCC4-/-细胞中提取的总RNA进行琼脂糖凝胶电泳和Ethylenediamine tetramine染色,以显示成熟的18S和28S rRNA。发现缺失METTL5或ZCCHC4基因的细胞正常产生成熟的rRNA。通过定量高分辨率Northern印迹进行的详细的pre-rRNA加工分析显示,所有主要的pre-rRNA种类均带有特异性探针,并且没有重大差异。总之,METTL5是18S rRNA m6A装饰酶,ZCCHC4 是28S rRNAm6A装饰酶,这两种酶都不是细胞生长或成熟rRNA产生所必需的,也不是任何沉积的修饰。
3.人METTL5与TRMT112形成异源二聚体,获得代谢稳定性
在作者最近的研究中,旨在描述古细菌中甲基转移酶激活剂Trm112的相互作用网络。发现METTL5同源基因HVO _1475(以下简称HVO Mettl5,35%的序列同一性METTL5),这导致测试HvoMettl5是否可以直接与HvoTrm112相互作用,并由此扩展人类METTL5是否可能与TRMT112相互作用。Hvo Mettl5与Hvo Trm112发生物理相互作用,而人类METTL5与TRMT112直接相关。当ZCCHC4与TRMT112共表达时,没有观察到稳定作用。通过X-射线衍射确定蛋白晶体结构。人TRMT112包含两个结构域:锌结合域(ZBD)和中央域。
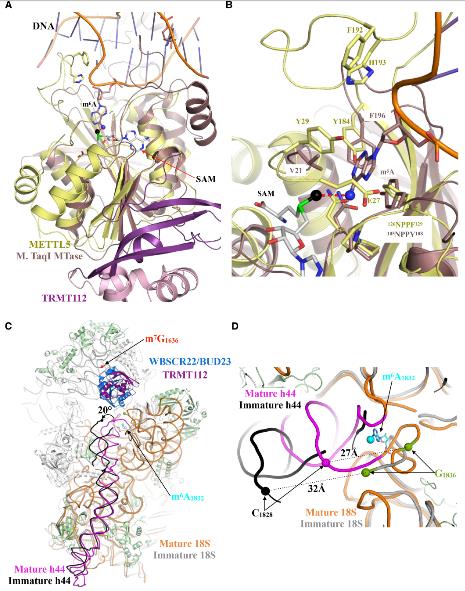
4.METTL5-TRMT112与DNA和RNAm6A甲基转移酶对核糖体生物发生的意义比较
METTL5-TRMT112的结构是继mElt3-METTL14,METL16和CAPAM之后的人类确定的第四个RNA m6A甲基转移酶结构。没有观察到METTL5-TRMT112与上述三种甲基转移酶有明显的相似性。这表明METTL5-TRMT112可能以其自己的特定方式运行。有趣的是,将METTL5-TRMT112的结构与结合到短双链DNA片段的m6A DNA甲基转移酶的结构进行比较时,发现了意想不到的共性。一个DNA链的目标腺嘌呤环从双螺旋中伸出并指向活性位点,并由105NPPY108标记以及Val21和Phe196侧链协调。这些分别对应于人METTL5的126NPPF129,Tyr29和Tyr184,这可能像M. TaqI一样在其活性位点目标为腺嘌呤,最佳地定位腺嘌呤以将甲基基团转移到其N6原子上。
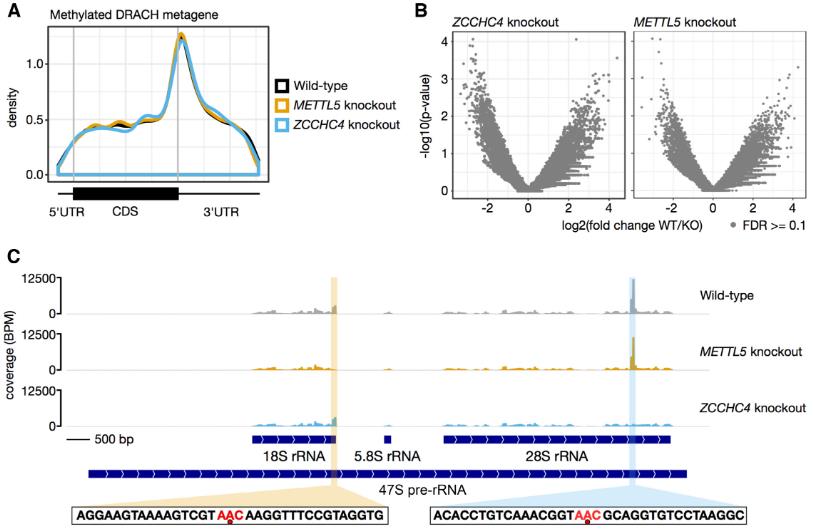
5.METTL5和ZCCHC4仅是m6A核糖体RNA的编辑器
我们想知道是否METTL5-TRMT112或ZCCHC4可以进行动作在核糖体RNA以外的RNA底物上。为了验证这一问题,使用miCLIP技术对缺少METTL5或ZCCHC4的人类细胞的转录组进行m6A的单核苷酸图谱。虽然12346个m6A位点重新定位在同基因对照细胞的mRNA上;在缺少METTL5或ZCCHC4的情况下,mRNA或lncRNA中的位点差异很大。miCLIP分析进一步证实,METTL5-TRMT112负责18S rRNA m6A 1832安装,ZCCHC4用于28S rRNA m6A 4220沉积。总之,METTL5和ZCCHC4分别对18S和28S rRNA m6A修饰具有高度特异性。
结论:
METTL5一个是负责18S rRNA m6A修饰的酶,并确认ZCCHC4为28S rRNAm6A修饰酶;METTL5必须与甲基转移酶激活剂TRMT112形成异源二聚体复合物,以在细胞中获得代谢稳定性。并且确定了METTL5-TRMT112的第一个原子分辨率结构。