






m6A甲基化与肝癌之间的奥秘
m6A甲基化与肝癌之间的奥秘
m6A甲基化是一种众所周知的具有表观遗传新功能的修饰,有报道称其参与了肝癌(HCC)的发生,为该疾病的分子发病机制提供了新的见解。然而,作为m6A甲基化的关键成分,WTAP在HCC中尚未得到很好的研究。在此,我们研究了WTAP在肝癌中的生物学作用及其机制.
本文利用组织芯片和TCGA数据集检测WTAP的表达及其与临床病理特征的关系。通过细胞增殖实验、菌落形成实验、Edu化验和皮下异种移植实验,阐明了WTAP对肝癌细胞的作用。另外,利用RNA测序结合GEO数据筛选WTAP候选靶点。最后,我们通过m6A点印记法、甲基化RNA免疫沉淀(MeRIP)、双荧光素酶报告基因检测、RNA免疫沉淀(RIP)和染色质免疫沉淀(ChIP)等方法研究了WTAP在HCC中的调控机制。
结果:
1. WTAP过表达与肝癌预后不良有关
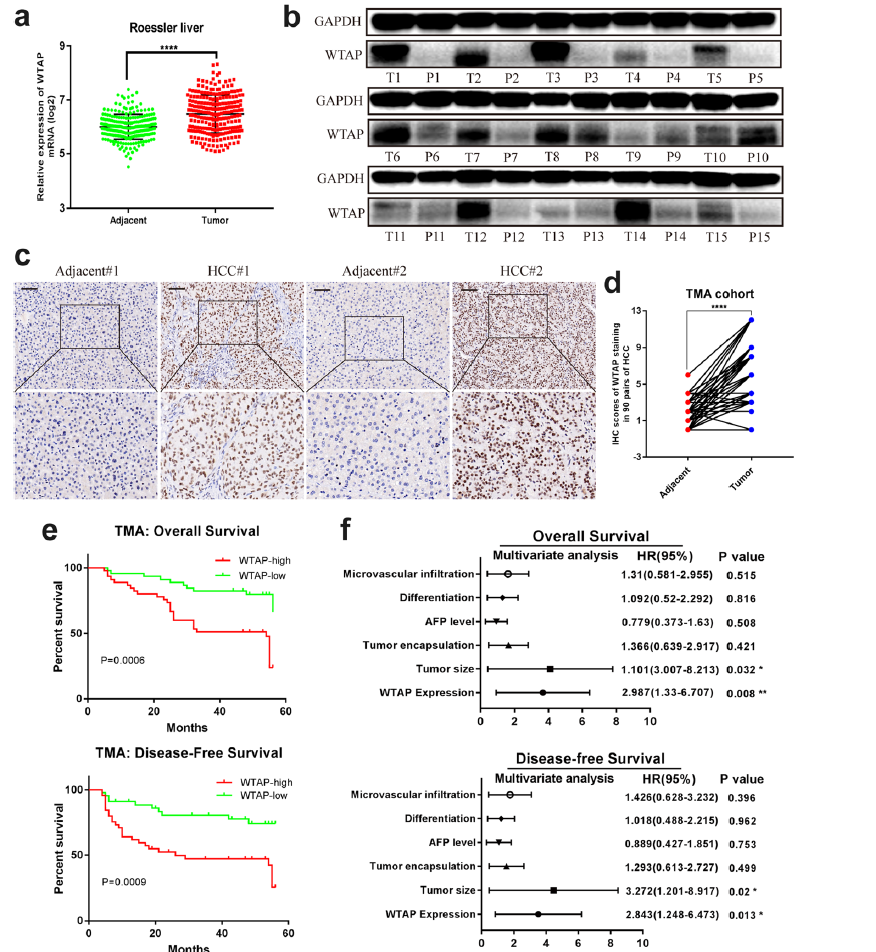
为了阐明WTAP的作用,我们首先从GEO数据集和TCGA数据中分析了WTAP在人HCC样本中的mRNA表达。结果显示WTAP在肿瘤组织中的表达显著增高(图1a)。WTAP蛋白水平在HCC组织中也明显上调(图1b),经TMA队列IHC染色进一步证实(图1c,d)。此外,探讨了90例HCC患者WTAP表达与临床病理特征的关系。Kaplan-Meier分析显示,WTAP表达水平越高的患者总体生存率(OS)和无病生存率(DFS)越低(图1e)。而且WTAP的过表达被发现是HCC患者的OS 和DFS 的独立预后因素(图1f)。综上所述,我们的结论是WTAP在HCC中上调,并与其不良预后密切相关。
2. WTAP在体内外均能促进肿瘤的增殖和致瘤能力
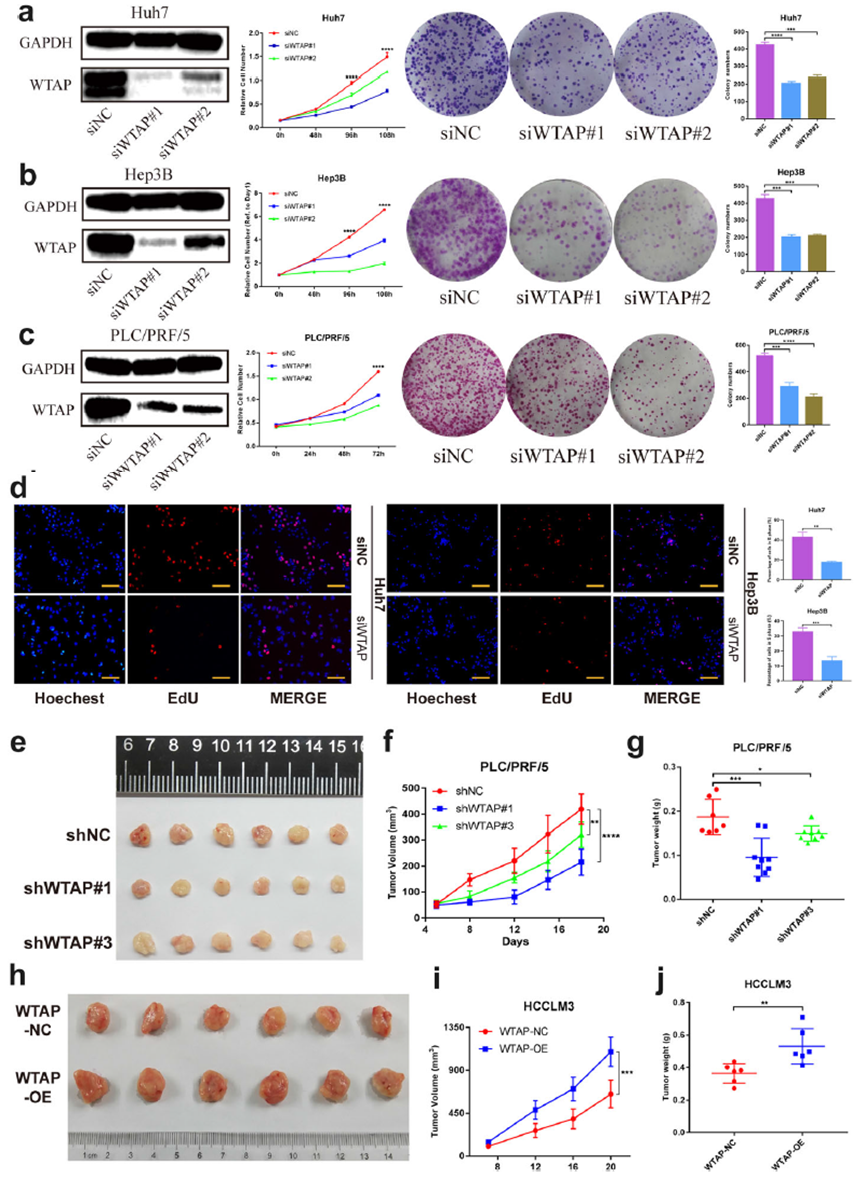
CCK-8和集落形成试验表明,WTAP缺乏抑制了三种细胞系(Huh7,Hep3B,PLC/PRF/5)的增殖能力(图2 a-c)。此外,EdU分析暗示敲低WTAP会减缓细胞生长(图2 d)。
为了证实WTAP在体内的作用,我们通过皮下注射HCC细胞,在裸鼠体内稳定敲低(shWTAP #1, #3)或过表达WTAP (WTAP- oe),构建了肿瘤异种移植模型。我们发现WTAP损耗抑制肿瘤发生(图2 e),显著降低肿瘤体积和重量(图2 f,g)。与此同时,WTAP的强迫表达导致异种移植小鼠的表型逆转(图2h-j)。综上所述,我们认为WTAP在HCC中具有肿瘤促进作用。
3.ETS1被认为是HCC中WTAP的潜在靶点
为了了解WTAP在HCC中发挥肿瘤促进作用的潜在机制,我们采用转录组测序来阐明WTAP敲除细胞的转录改变。WTAP缺失后,层次聚类显示上调基因1636个,下调基因794个(图3a)。为了缩小下游靶点的范围,我们整合Liu等人发表的GEO数据(GSE46705)。通过MeRIP -seq和CLIP-seq对WTAP调控的转录本进行精化,建立基因集重叠。维恩图共揭示了15个基因(图3b)。然后我们进行RT-qPCR来评估WTAP对每个候选基因的影响。其中,在WTAP沉默后,ETS1和ETS2持续显著上调,当WTAP过表达时,ETS1和ETS2均适度下调(图3c-e)。然而,western blot分析显示,WTAP失活明显导致ETS1蛋白水平升高,而不是ETS2蛋白水平升高(图3f,g)。接着我们进一步证明,在Hep3B和HCCLM3细胞中,WTAP对ETS1的含量有相反的影响(图3h,i)。因此,我们推测ETS1功能障碍可能是WTAP介导的HCC增殖特征的原因。

4. WTAP增加了ETS1 mRNA的m6A修饰,并以m6A-HuR介导的方式抑制ETS1的表达
为了确定ETS1的m6A修饰是否通过WTAP介导,我们首先通过两种不同的方法(m6A斑点印迹法和RNA甲基化定量法)检测阴性对照组和稳定的WTAP敲低组的m6A的整体水平。结果表明,随着WTAP的缺失,两株HCC细胞系m6A水平显著降低(图4a,b)。通过MeRIP-qPCR检测m6A在ETS1中的富集情况。发现与IgG对照相比,m6A特异性抗体较强地富集了ETS1转录本。此外,我们发现在WTAP沉默后,m6A修饰的ETS1显著减少(图4c)。因此,我们认为WTAP能够影响m6A的总体水平,尤其是ETS1。
为了证实m6A修饰ETS1的要求,我们用野生型(WT)和两个突变体(Mut)质粒进行了荧光素酶报告基因测定。对于Mut报告基因,预测m6A位点的几个腺苷碱基(A)被胞嘧啶碱基(C)取代,以消除m6A甲基化的影响,而WT报告基因包含完整的m6A位点(图4d)。正如所料,WT组的荧光素酶活性在WTAP敲低作用下适度增强,而Mut组的荧光素酶活性对WTAP沉默的影响具有一定的抵抗作用(图4e),说明ETS1的调控受WTAP诱导的m6A修饰的控制。总之,这些数据表明WTAP足以调控ETS1 mRNA的m6A修饰。
RNA结合蛋白HuR易于与较少的m6A修饰的RNA结合,并稳定其结合的对应物,因此我们认为HuR可能调控ETS1。结果表明,HuR的减少显著减轻了ETS1的表达(图4f)。我们发现,与IgG对照组相比,HuR特异性抗体显著富集ETS1 mRNA,而WTAP的抑制显著增强了ETS1 mRNA的富集(图4g)。此外,WTAP敲低引起的ETS1升高可以通过HuR的衰减来恢复(图4h)。我们还发现,敲除WTAP可以延长ETS1 RNA的半衰期,而HuR沉默可以逆转这种效果(图4i)。最后,我们还进行了荧光素酶报告基因测定,以确定HuR参与m6A的修饰。在WT组中,HuR可以挽救WTAP沉默引起的荧光素酶活性的增强。然而,当ETS1的m6A位点可能发生突变时,HuR表达的改变似乎对荧光素酶活性没有影响(图4j)。总之,我们的发现表明WTAP通过m6A- HuR依赖的通路抑制ETS1。


5. ETS1作为一种肿瘤抑制因子,逆转了WTAP在HCC中的作用
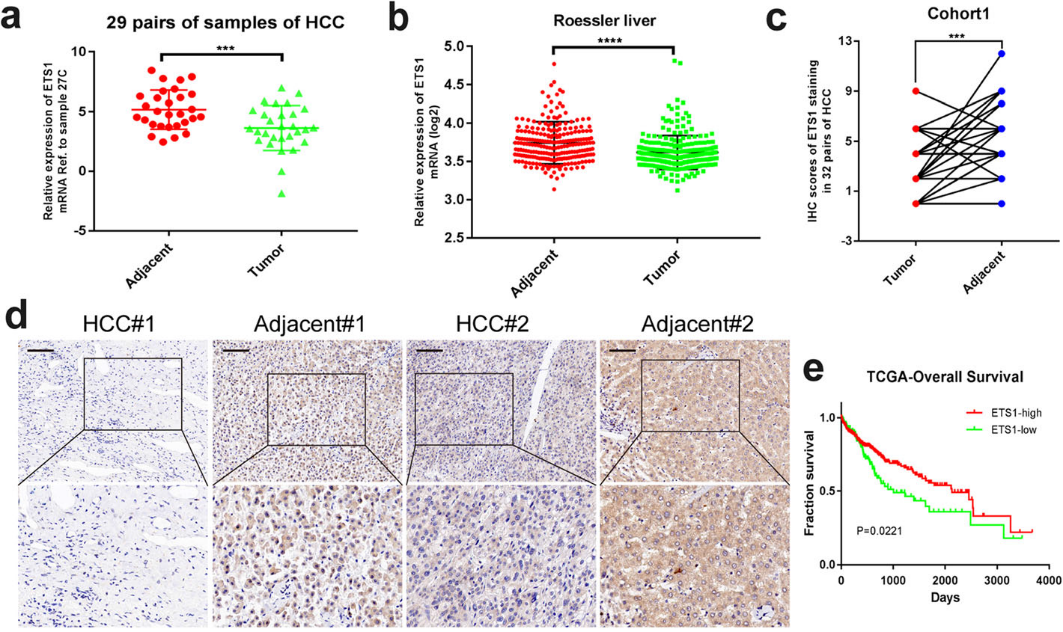
与癌旁组织相比,肿瘤组织中ETS1的表达较低(图5a,b)。通过对cohort1的免疫组化染色,我们发现邻近组织的阳性率明显较高(图5c,d)。根据TCGA数据,ETS1水平越高,预后越好(图5e)。体外功能验证表明,ETS1的敲除增强了MHCC97H、HCCLM3和Huh7细胞的增殖能力(图5f,g)。此外,ETS1沉默足以挽救WTAP敲低对HCC细胞生长和存活的抑制作用(图5h,i),因此ETS1的失活可能通过WTAP-ETS1轴导致HCC的进展。

6. WTAP的沉默通过ETS1-p21/p27轴在HCC中引起G2/M阻滞
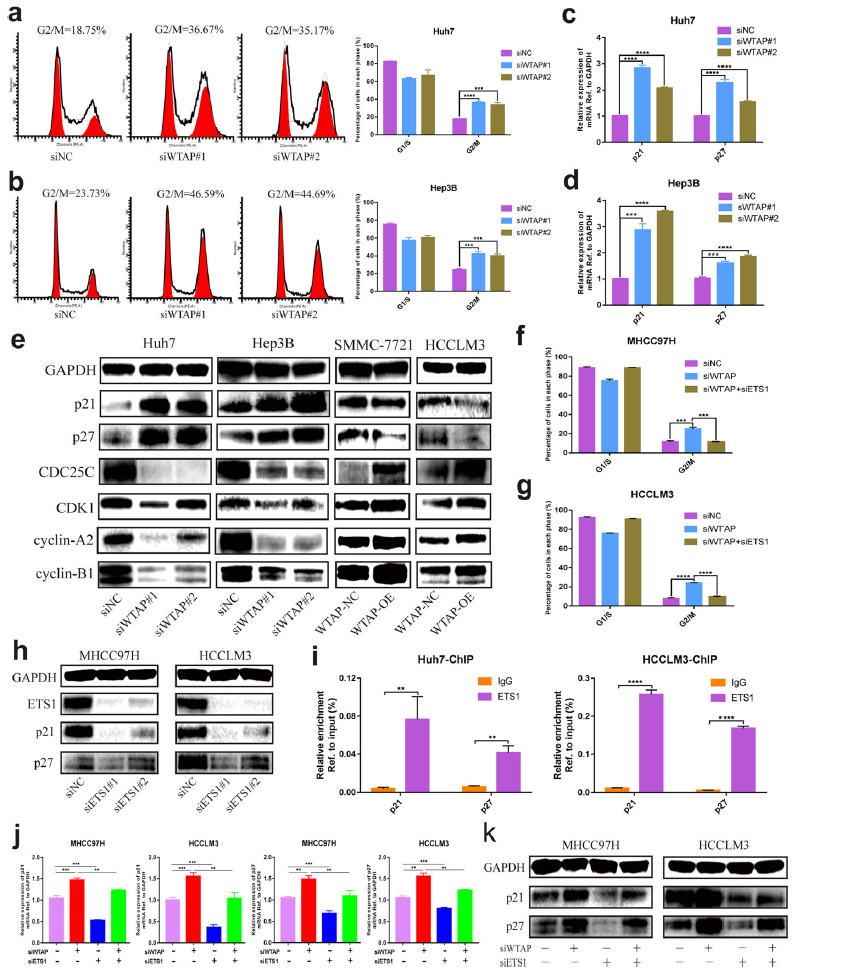
据报道WTAP参与细胞周期调控,因此我们拟探讨其在HCC中的相关机制。结果表明,WTAP敲低导致了G2/M的大量阻滞(图6a,b)。为了阐明潜在的机制,我们研究了与细胞周期相关的几个元素。有趣的是,当WTAP在转录水平失活时,在细胞周期和增殖中发挥重要作用的肿瘤抑制因子p21和p27的表达显著增加(图6c、d)。同时,western blot分析发现WTAP敲低上调了p21和p27,同时下调了CDC25C、CDK1、cyclin-A2和cyclin-B1。相反,WTAP的过表达减轻了p21和p27的表达,降低了G2期所有指示的检查点蛋白的放大(图6e)。
为了进一步研究ETS1是否参与WTAP诱导的细胞周期效应,我们进行了一系列的挽救性实验。结果表明抑制ETS1可显著逆转WTAP沉默引起的G2/M阻滞(图6f)。此外,ETS1的下调导致p21和p27的减少(图6h)。而且我们推测ETS1可能增强p21和p27的转录,因为ChIP实验表明ETS1可以与启动子结合(图6i)。此外,ETS1的抑制恢复了WTAP抑制引起的p21和p27表达升高(图6j,k),这些数据表明ETS1-p21/p27轴在HCC中对WTAP依赖的细胞周期调控至关重要。
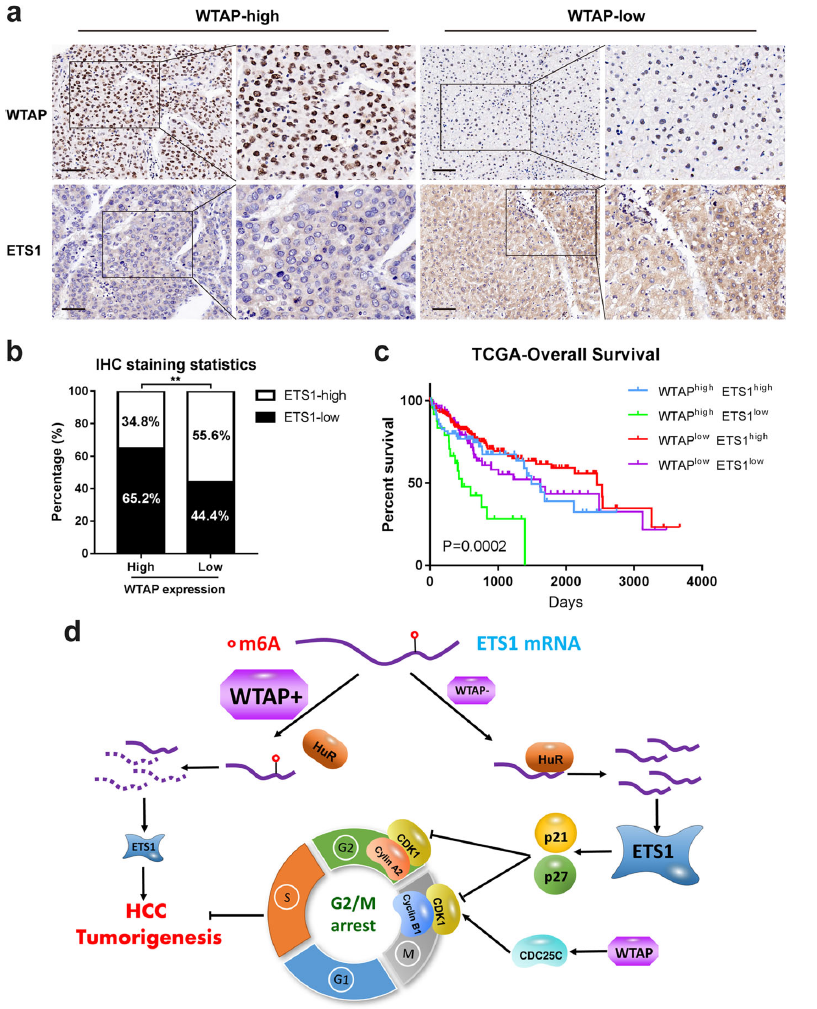
7.结合高WTAP和低ETS1表达预测肝癌的不良预后
为了评估WTAP和ETS1的临床相关性,我们对来自cohort1的相同HCC标本进行了WTAP和ETS1染色的IHC检测。近65.2%的高WTAP表达的样本ETS1染色较弱,而大约55.6%的低WTAP表达的样本表现出更强的ETS1染色(图6a,b)。基于这两种因素结合的Kaplan-Meier分析进一步证明,WTAPhighETS1low表达的HCC患者的总体生存率比其他任何组都要低(图7c)。综上所述,WTAP与ETS1在临床样本中呈负相关关系,WTAP与ETS1的共表达模式可能是判断HCC预后的一个有效因素。
总结:
总之,我们的研究阐明了WTAP和活化的WTAP介导的m6A机制在HCC中的致癌作用。WTAP上调参与了ETS1的m6A修饰,随后通过HuR关联的方式使ETS1表观遗传沉默。我们的发现丰富了肿瘤特征中m6A甲基化的功能价值,为探索肝癌治疗中有效的治疗策略开辟了潜在的途径。