






外泌体与糖尿病伤口愈合
目前,开发改善糖尿病个体伤口愈合的治疗策略仍然具有挑战性。外泌体代表了一种有前途的纳米材料,可以从中分离出microRNA(miRNA)。这些miRNA具有发挥治疗作用的潜力,因此,确定外泌体中循环的特定miRNA的潜在治疗作用非常重要。在这里,小编与大家分享一篇近期发表于Small上的文章“Circulating Exosomal miR-20b-5p Inhibition Restores Wnt9b Signaling and Reverses Diabetes-Associated Impaired Wound Healing”。在本研究中,我们从2型糖尿病患者或健康对照组中分离出外泌体,并评估了其在糖尿病伤口愈合中的作用。

结果:
1.来自患有或不患有2型糖尿病的志愿者的外泌体特征
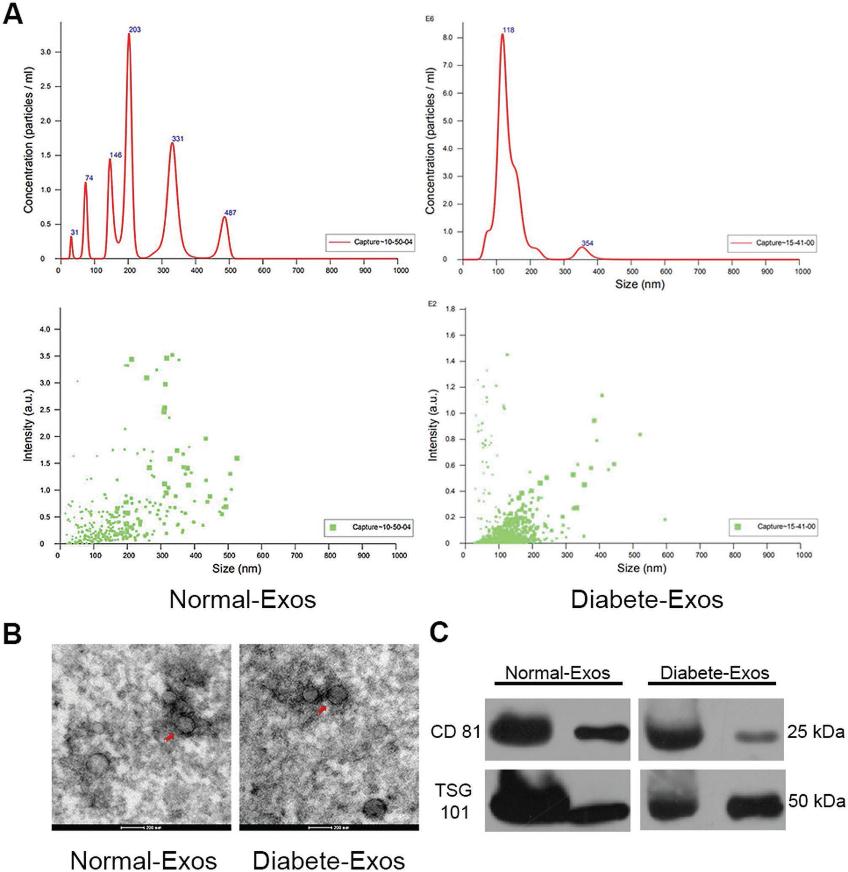
我们首先通过透射电子显微镜(TEM),动态光散射(DLS)和WB评估从志愿者患者血浆中分离的外泌体。DLS揭示了外泌体大小在30到200 nm之间,与先前有关外泌体大小的报道一致(图1A)。透射电镜显示这些颗粒具有杯状或球形的形态(图1B)。[12] WB进一步证实,这些颗粒包含外泌体表面标记,例如CD81和TSG101(图1C)。 这些结果一起证实了分离的循环纳米颗粒是外泌体。
2.来自2型糖尿病患者的外泌体阻碍小鼠创面愈合
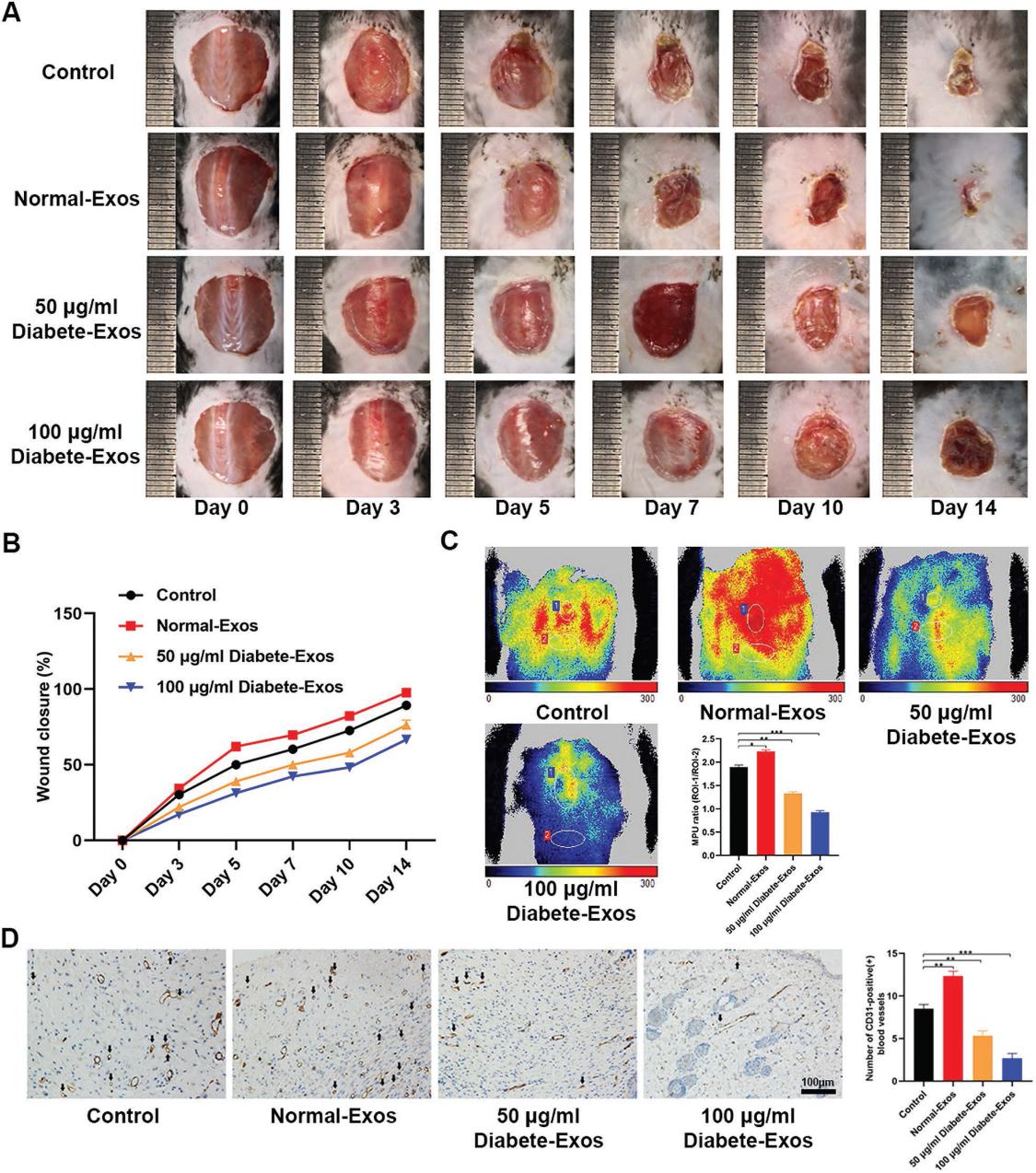
我们探索了来自2型糖尿病患者的外泌体对伤口愈合能力的影响。我们观察到注射糖尿病患者的外泌体时伤口闭合速率明显降低(图2A,B),与非糖尿病外泌体对照相比,治疗后10天伤口部位的平均灌注单位(MPUs)比率更低(图2C) 。 此外,我们量化了伤口部位的CD31表达,进一步证实了糖尿病患者的外泌体能够抑制局部血管生成(图2D)。 这些发现共同表明,糖尿病患者的外泌体能够延迟体内伤口愈合。
3.来自2型糖尿病患者的外泌体抑制人脐静脉内皮细胞(HUVEC)增殖和血管形成并促进细胞凋亡
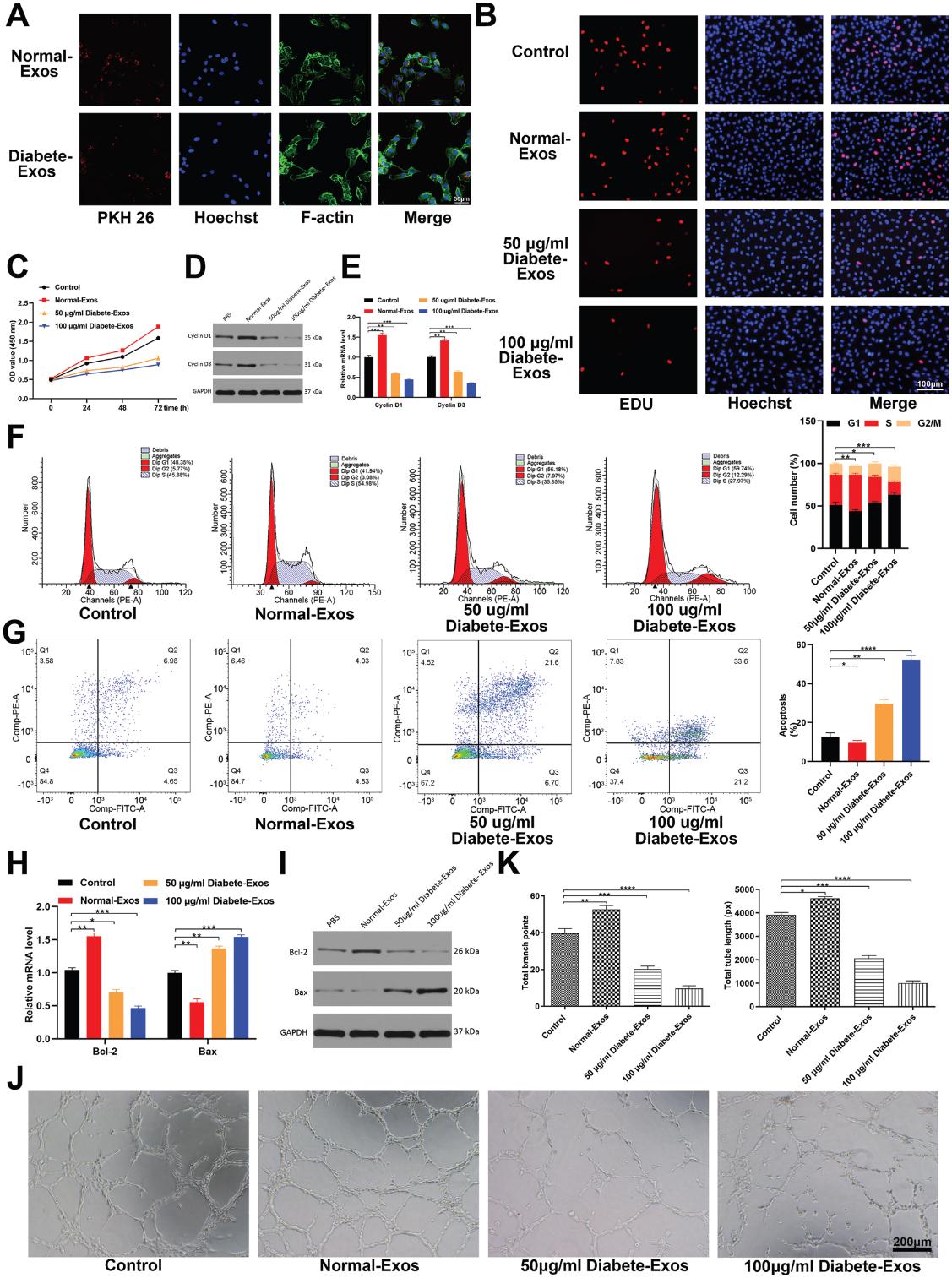
我们通过用PKH26染料标记外泌体并将其添加到HUVEC培养基中来评估HUVEC在体外吸收循环外泌体的能力。结果显示,基于细胞中的红色荧光染色,HUVECs能够在12小时内吸收非糖尿病和糖尿病患者的外泌体(图3A)。接下来,我们使用CCK-8和EDU掺入法评估了HUVEC增殖,从而探讨了糖尿病患者的外泌体对HUVEC功能的影响。 我们发现,相对于非糖尿病患者的外泌体治疗,用50-100μg mL-1糖尿病患者的外泌体治疗HUVEC足以以剂量依赖性方式显著降低HUVEC增殖(图3B,C)。 我们进一步发现,在糖尿病患者的外泌体治疗的HUVEC中,与增殖相关的蛋白Cyclin D1和Cyclin D3减少了(图3D,E)。 流式细胞术进一步定量处理细胞的细胞周期分布和凋亡率,发现糖尿病患者的外泌体处理后进入S期的细胞比例明显下降,而HUVEC凋亡相应增加(图3F,G)。 此外,我们观察到糖尿病患者的外泌体治疗后Bcl-2表达降低,而Bax表达升高(图3H,I)。最后,我们探索了糖尿病外泌体调节HUVEC血管生成的能力,并发现糖尿病患者的外泌体治疗后管形成显著减少(图3J,K)。 总之,这些发现表明来自2型糖尿病患者的外泌体会损害体外伤口的愈合。
4.2型糖尿病外泌体含有高水平的miR-20b-5p并将此miRNA转移到HUVECs
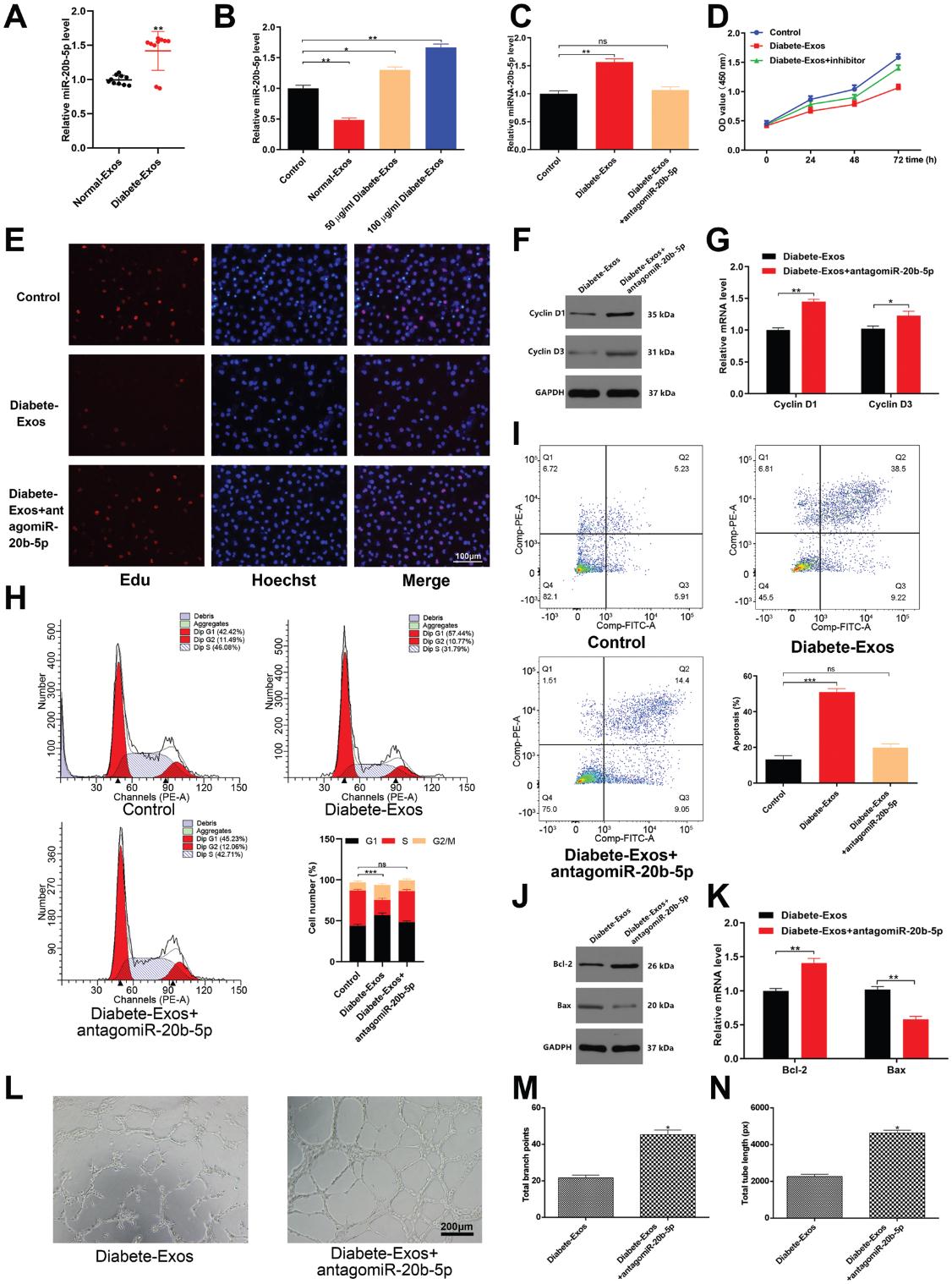
在先前的研究中,已发现miR-20b-5p在2型糖尿病患者的外泌体中高表达,同样,我们证实了与非糖尿病对照组相比,糖尿病患者的外泌体中该miRNA的水平升高(图4A )。 迄今为止,我们已经表明糖尿病患者的外泌体可以被HUVEC吸收,接下来我们评估包裹在这些外泌体中的miRNA是否可以被类似地吸收并在HUVEC中保持功能。与这种功能一致,我们发现相对于非糖尿病患者的外泌体治疗的HUVEC,miR-20b-5p水平在糖尿病患者的外泌体治疗的HUVEC中有显著增加(图4B)。为了探索miR-20b-5p介导糖尿病患者的外泌体对HUVEC功能的影响,我们用miR-20b-5p抑制剂(antagomiR-20b-5p)转染了这些外泌体处理的细胞,结果显示antagomiR-20b-5p转染后miR-20b-5p表达下降(图4C)。 CCK8和EDU分析证实,如果还用antagomiR-20b-5p处理,则用糖尿病患者的外泌体处理的细胞的增殖也会增强(图4D,E)。 AntagomiR-20b-5p转染还导致Cyclin D1和Cyclin D3表达增加,以及在S期的HUVEC百分比更高(图4F,H)。我们同样证实了antagomiR-20b-5p处理降低了糖尿病患者的外泌体诱导的HUVEC凋亡的能力(图4K),并且在antagomiR-20b-5p处理的细胞中,血管的形成活性也得到了类似的增强(图4L,N)。总之,这些结果表明,糖尿病患者的外泌体中富含miR-20b-5p,可以调节HUVEC功能。
5.miR-20b-5p抑制HUVECs的血管生成作用
我们探索了miR-20b-5p通过使用agomiR或agomiR-20b-5p处理细胞而直接影响HUVEC的能力,后者显著提高了miR-20b-5p的表达(图5A)。 处理后,CCK8和EDU结果证实了miR-20b-5p抑制HUVEC增殖(图5B,C)并降低细胞周期蛋白D1和D3表达的能力(图5D,E)。 经agomiR-20b-5p处理后,HUVECs更多地处于G1期和凋亡(图5F,G),而用该miRNA转染后,Bax表达增加而Bcl-2表达降低(图5H,I)。 同样,miR-20b-5p抑制了HUVEC管的形成(图5J L)。这些结果证实了miR-20b-5p在体外直接抑制HUVEC功能的能力。
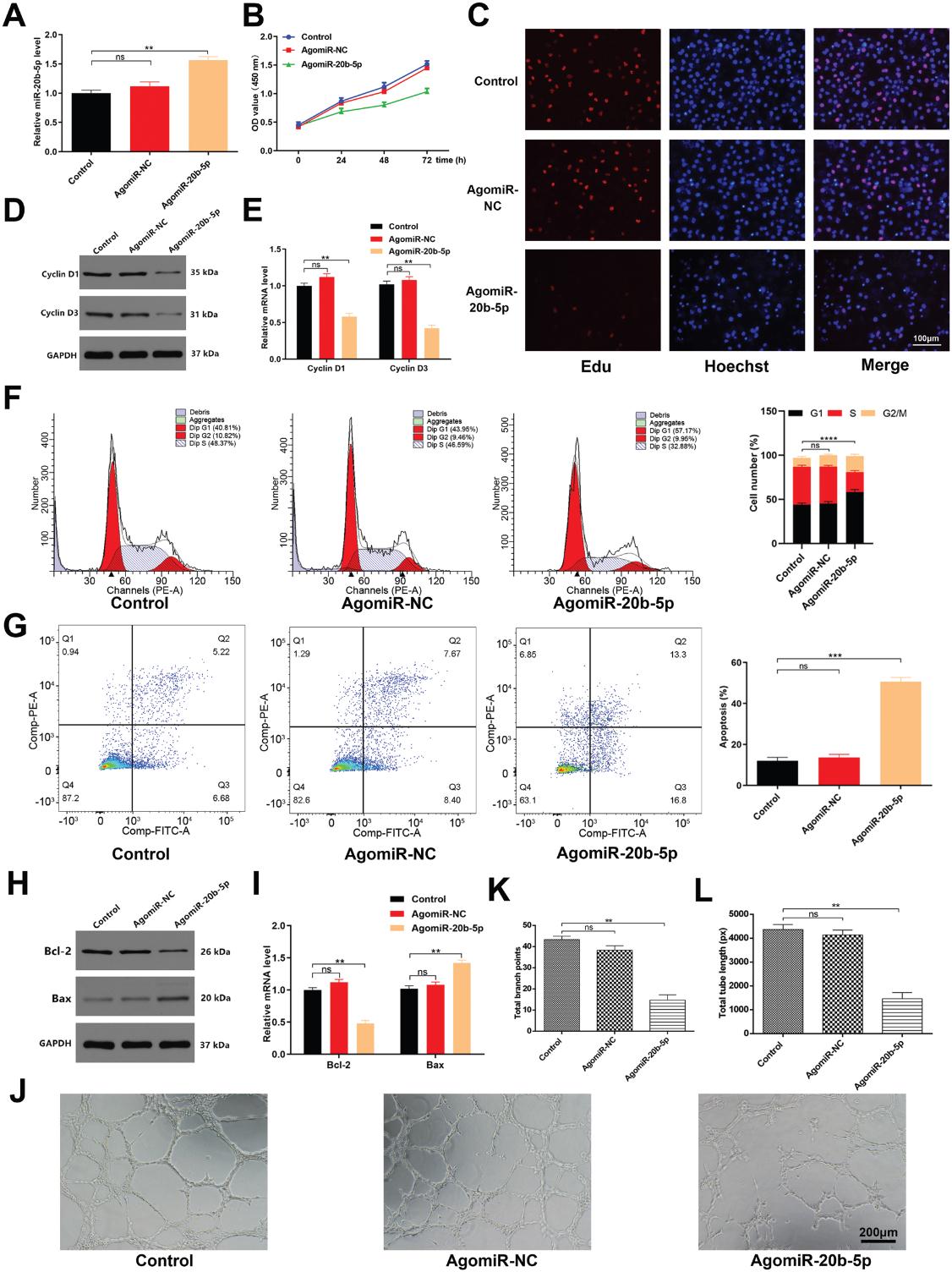
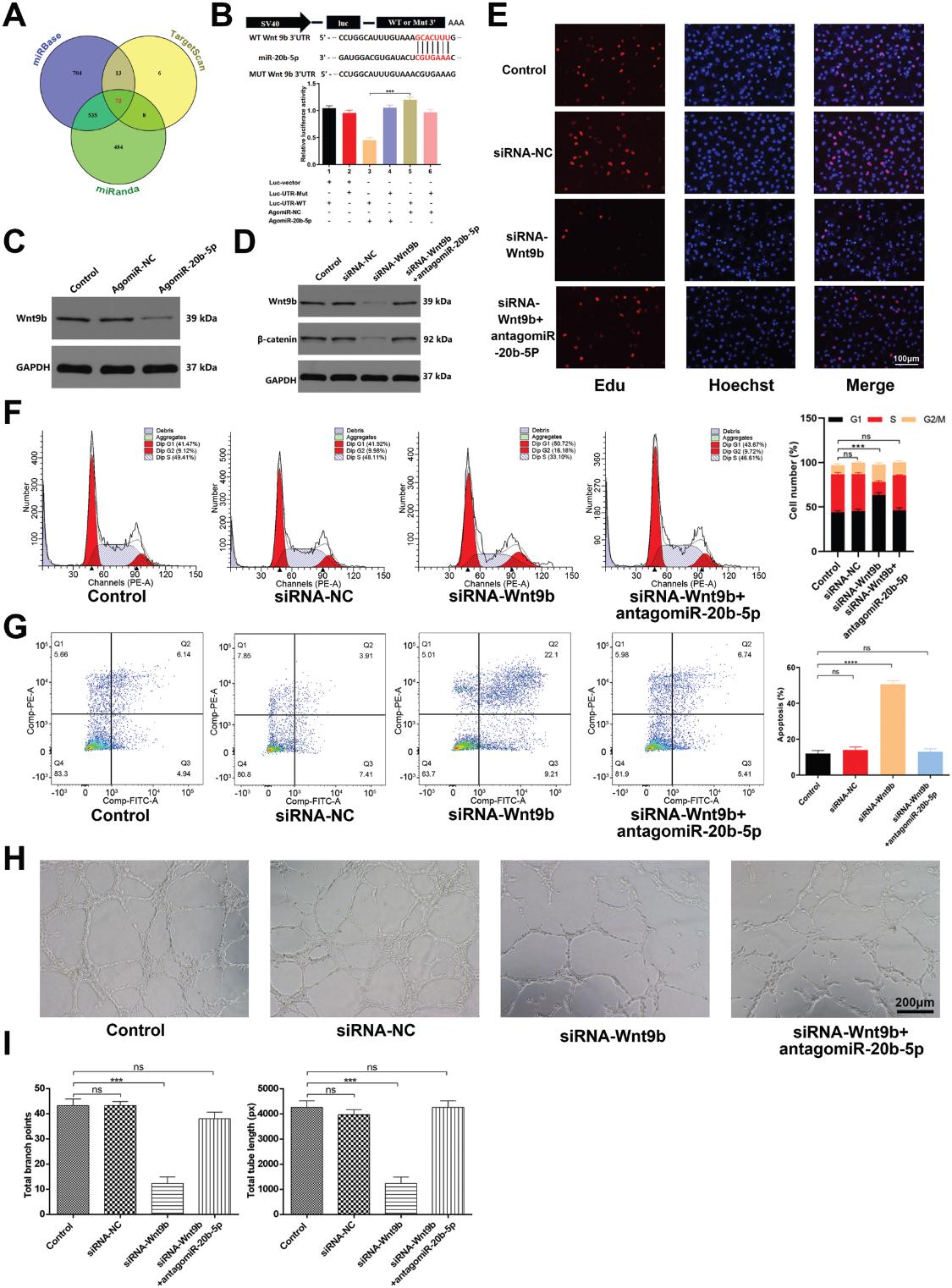
6.miR-20b-5p发挥抗血管新生的影响通过抑制Wnt9b /β-Catenin通路
我们评估了miR-20b-5p抑制HUVEC功能的分子机制。为此,我们试图使用miRbase,Targetscan和miRanda鉴定推定的miR-20b-5p靶标,从而使我们将Wnt9b鉴定为值得验证的靶标基因。如图6A所示,鉴定出73个基因为miR-20b-5p靶基因。先前的研究报道血管生成与wnt /β-catenin信号通路密切相关。在仔细筛选了73个基因后,我们发现Wnt9b可能是参与miR-20b-5p介导的血管生成的最有潜力的基因。此外,我们发现miR-20b-5p过表达显著抑制Wnt9b的表达的(图6B)。我们通过荧光素酶报告基因实验证实miR-20b-5p能够特异性结合到Wnt9b mRNA的预测靶区。当这个靶区发生突变时,miRNA不再能够结合抑制荧光素酶活性(图6C)。为了证实HUVEC血管生成依赖于Wnt9b,我们使用针对Wnt9b的siRNA转染HUVECs。该siRNA显著降低了处理过的细胞中Wnt9b和β-catenin的表达(图6D),抑制了细胞增殖和管形成,同时促进了细胞凋亡(图6E-I)。抑制miR-20b-5p活性部分逆转了siRNA-Wnt9b对HUVECs的抗血管生成作用。这些发现支持miR-20b-5p直接靶向Wnt9b的能力,从而抑制HUVEC的血管生成,因此表明该miRNA至少部分地通过调节Wnt9b/β-catenin 信号通路起作用。
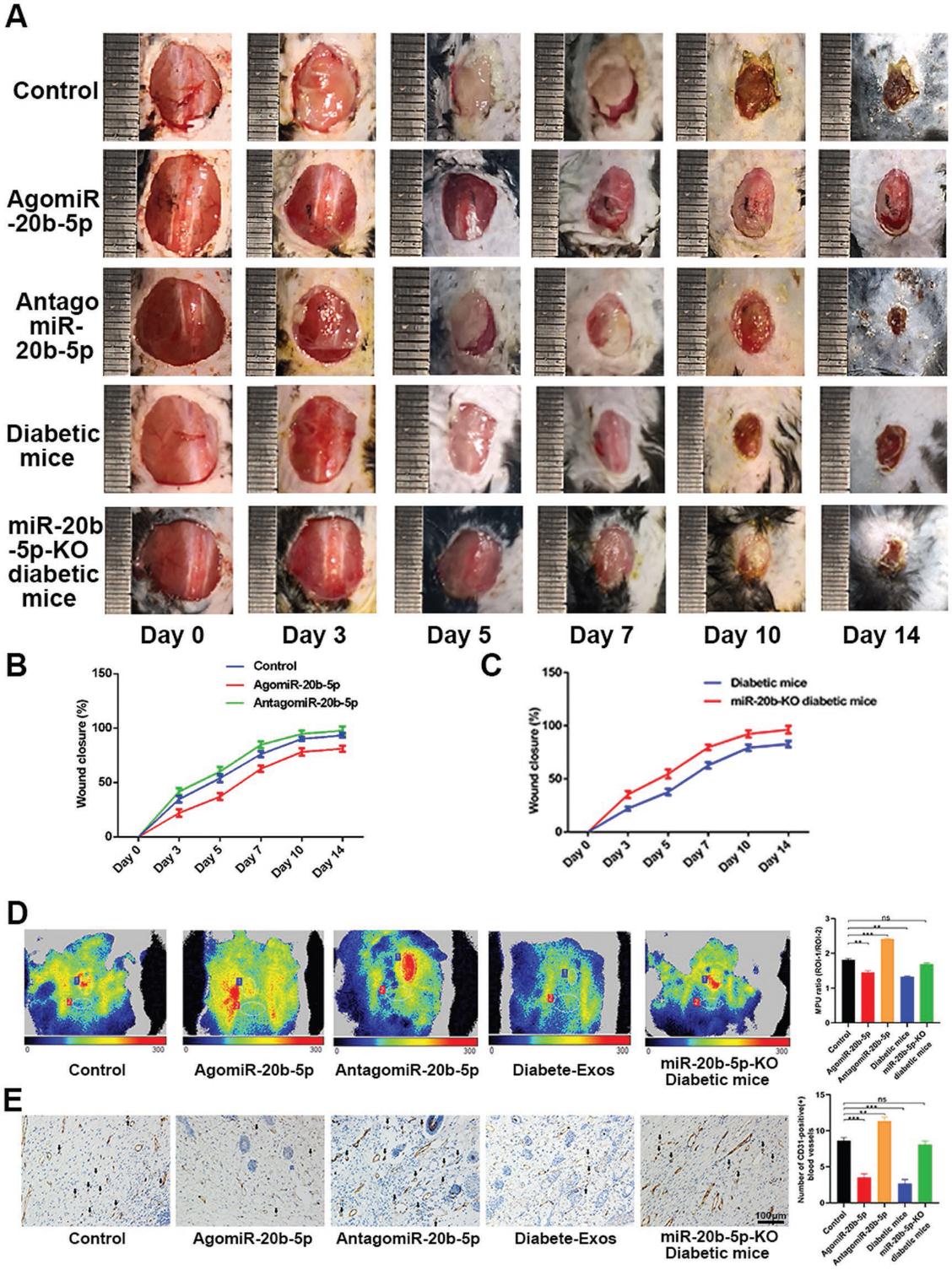
7.miR-20b-5p在体内延迟伤口愈合
最后,我们在0、3、5、7、9和11天局部注射PBS、agomiR-20b-5p或antagomiR-20b-5p来评估miR-20b-5p抑制体内伤口愈合的能力。我们发现局部的agomiR-20b-5p注射显著损害了伤口的愈合,局部MPU比值降低与miRNA延迟伤口修复的能力一致。 为了进一步探索这种miRNA如何影响伤口愈合,我们在WT或miR-20b KO糖尿病模型小鼠的背部产生了伤口,并在miR-20b KO糖尿病小鼠中观察到了优异的伤口愈合(图7A-E)。因此这些发现表明miR-20b-5p沉默能促进伤口愈合,从而突出其作为糖尿病个体治疗靶点的潜力。
结论:
我们的体外和体内研究结果均表明,糖尿病患者血浆外泌体通过在体外和体内影响血管内皮细胞的血管生成活性来抑制皮肤伤口愈合。miR-20b-5p在外泌体中的富集通过抑制Wnt9b /β-catenin信号通路在损害伤口愈合中起重要作用。 因此,将纳米材料与miR-20b-5p抑制剂结合使用可能是将来促进糖尿病伤口愈合的有前途的治疗方法。