






m6A阅读器YTHDF1促进卵巢癌的进展
m6A是哺乳动物mRNA中最丰富的RNA修饰,越来越多的证据表明m6A在人类肿瘤发生中起着关键作用。然而,m6A的阅读器YTHDF1,是否针对一个参与蛋白质翻译的基因,从而影响癌细胞的整体蛋白质生产,目前还没有很多的研究。因此小编为大家详细介绍发表于“Nucleic Acids Research”杂志的文章“The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation”。在这里,通过对卵巢癌的多组分分析,我们确定了一种新的机制。YTHDF1通过与m6A修饰的EIF3C mRNA结合以m6A依赖的方式增强EIF3C的翻译,同时促进整体翻译输出,从而促进卵巢癌的发生和转移。

结果:
1.YTHDF1在卵巢癌中高表达
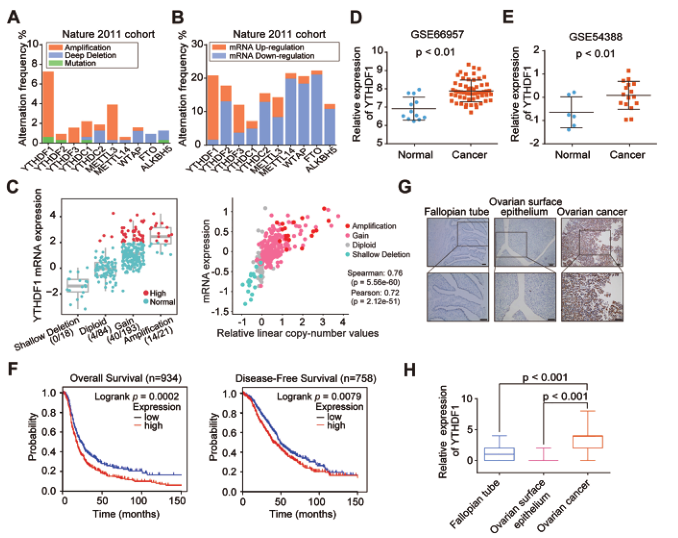
为了研究m6A相关基因在卵巢癌发展中的作用,我们首先利用cBioPortal TCGA数据集分析这些基因的遗传改变和表达水平。其中YTHDF1基因最常被扩增,其表达显著升高,并且YTHDF1的拷贝数状态与其在所有三个队列中的mRNA表达呈正相关(图1A-C)。与正常卵巢上皮细胞相比,YTHDF1在卵巢癌中表达上调(图1D和E)。此外,我们进行了Kaplan-Meier生存分析,发现高表达YTHDF1的卵巢癌患者总体生存率和无病生存率较差(图1F)。为了更准确地检测YTHDF1在卵巢癌中的表达,我们进行了组织芯片分析。结果显示,YTHDF1蛋白在卵巢癌样本中的表达高度增强(图1G和H)。同时,这些结果提示m6A读卡器YTHDF1在卵巢癌中高表达,并与卵巢癌患者的不良预后相关。
2.YTHDF1调节卵巢癌细胞的增殖、迁移和侵袭
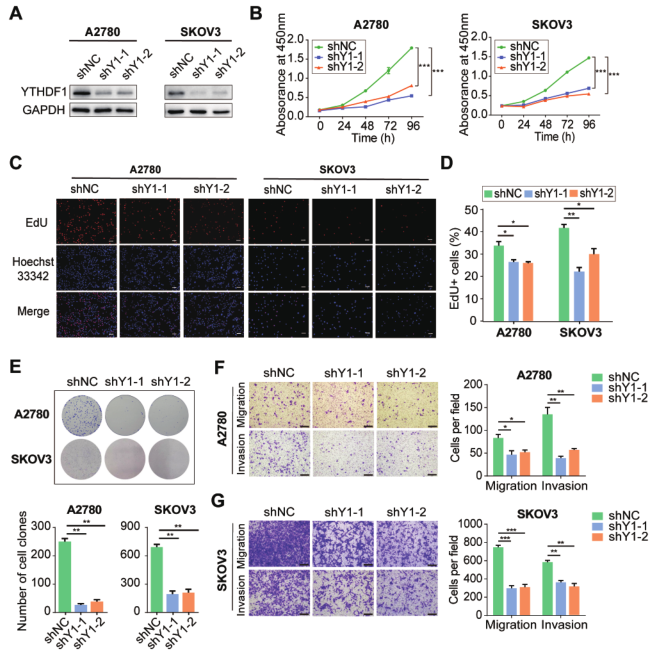
为了探讨YTHDF1在卵巢癌中的作用,我们试图对缺失YTHDF1的卵巢癌细胞的表型改变进行研究。shY1-1和shY1-2在A2780和SKOV3卵巢癌细胞中有效地击倒了YTHDF1(图2A)。通过CCK8检测,YTHDF1基因敲除显著损害A2780和SKOV3细胞的生长(图2B)。此外,EdU染色显示YTHDF1沉默后卵巢癌细胞的增殖显著降低(图2C和D)。菌落形成分析显示,YTHDF1基因敲除后克隆形成能力受到抑制(图2E)。此外,细胞迁移和侵袭分析显示,YTHDF1缺陷损害了A2780和SKOV3细胞的迁移和侵袭能力(图2F和G)。提示YTHDF1在卵巢癌细胞的增殖、迁移和侵袭中起重要作用。
3.YTHDF1缺乏抑制卵巢癌细胞在体内的肿瘤发生和转移
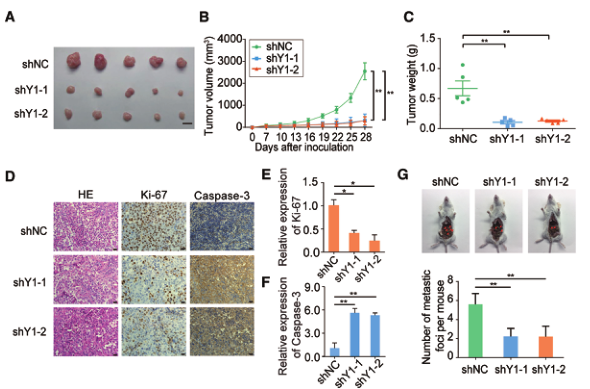
为了评价YTHDF1在卵巢癌体内的致癌作用,我们采用皮下肿瘤模型和腹膜转移异种移植模型。首先,在裸鼠皮下分别注射YTHDF1缺陷的A2780卵巢癌细胞和对照细胞。研究表明,来自YTHDF1缺陷组的肿瘤明显小于来自对照组的肿瘤(图3A)。同时,与对照组相比,YTHDF1基因敲除小鼠的平均肿瘤体积和牺牲时的体重显著降低(图3B和C)。我们还评估了这些实体瘤的细胞增殖和凋亡指数。与对照细胞相比,YTHDF1沉默的肿瘤显示出Ki-67信号降低,但caspase-3活性增加(图3D-F)。注射YTHDF1沉默的细胞实质上消除了细胞在腹腔内形成二次肿瘤的能力(图3G)。总之,这些结果显示了YTHDF1通过调节细胞增殖和转移在卵巢癌中的致癌作用。
4.卵巢癌YTHDF1靶点的鉴定
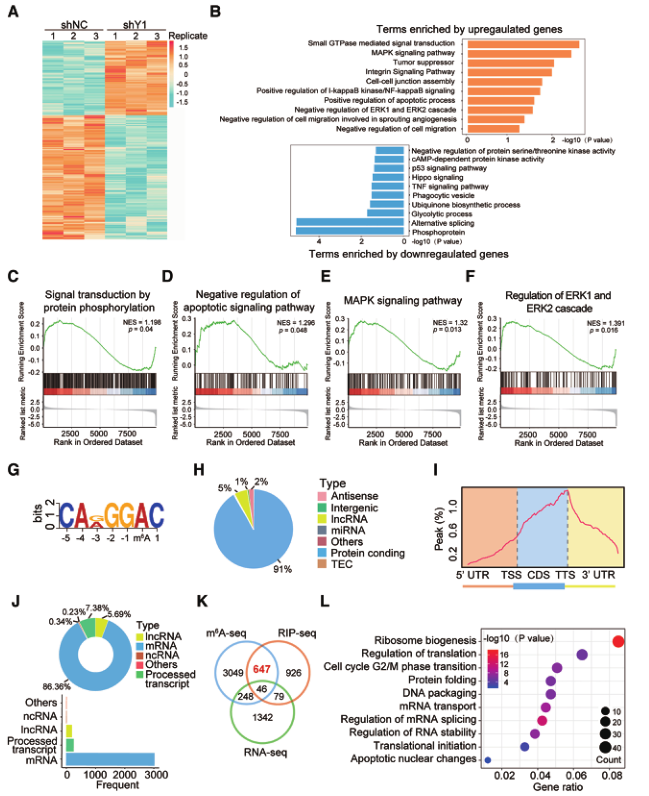
为了探讨YTHDF1在卵巢癌发生发展中的作用机制,我们对YTHDF1基因敲除后的A2780细胞进行了RNA序列分析。YTHDF1缺失导致1715个基因发生改变,包括633个上调基因和1082个下调基因(图4A)。MAPK信号途径、肿瘤抑制和细胞迁移调控多种富集途径和基因集富集分析(GSEA)揭示了YTHDF1基因的改变与蛋白质磷酸化、凋亡信号途径、MAPK途径、ERK级联和细胞活化等信号转导有关(图4B-F),支持YTHDF1在卵巢癌发生中的调节作用。
YTHDF1起着m6A阅读器的作用,通过结合和影响m6A甲基化转录本发挥作用。我们分析确定了m6A共有基序(GGAC),表明m6A修饰的mRNA成功富集(图4G)。这些m6A修饰主要位于蛋白质编码转录本中,并在终止密码子附近富集(图4H 和4 I)。RIP-seq揭示了1698个潜在的YTHDF1候选靶点,其中93%是mRNA,这些基因在不同的途径中富集(图4J)。有趣的是,来自RNA-seq、m6A-seq和RIP-seq的基因重叠显示,693个由YTHDF1结合的基因被m6A标记,其中647个(93.4%)基因在YTHDF1敲除后没有改变(图4K)。这些结果表明,YTHDF1并不影响其靶点的RNA丰度。此外,功能注释显示,这647个基因参与了包括翻译、mRNA剪接和RNA内稳态在内的多个RNA代谢过程(图4L)。
5.EIF3C是YTHDF1的m6A修改靶点
为了全面探讨YTHDF1与其靶向转录本之间的直接相互作用,我们进行了eCLIP-seq,鉴定了2343个靶向转录物,这些转录物大部分是mRNAs(图5A)。从eCLIP seq中还鉴定出m6A一致基序GGAC和YTHDF1结合偏好(图5B和5C)。值得注意的是,175个基因被确认为YTHDF1的直接靶点(图5D)。Circos图显示来自RIP-seq或eCLIP-seq的YTHDF1靶向基因被m6A修饰,但是它们的转录在YTHDF1基因敲除后没有改变(图5E),这表明YTHDF1可能调节卵巢癌细胞的翻译而不是RNA丰度。
接下来,我们分析了175个YTHDF1结合的mRNA中m6A峰和eCLIP峰的分布,其中一些与肿瘤的发生密切相关。我们发现这些转录本中的大多数YTHDF1结合位点,包括EIF3C,与m6A 位点吻合良好(图5F-J)。GO分析显示,YTHDF1结合转录本与翻译调控关系最为密切(图4L)。CLIP qPCR显示,YTHDF1最显著地富集了EIF3C mRNA(图5K)。这些数据表明EIF3C是卵巢癌细胞YTHDF1的直接靶点。
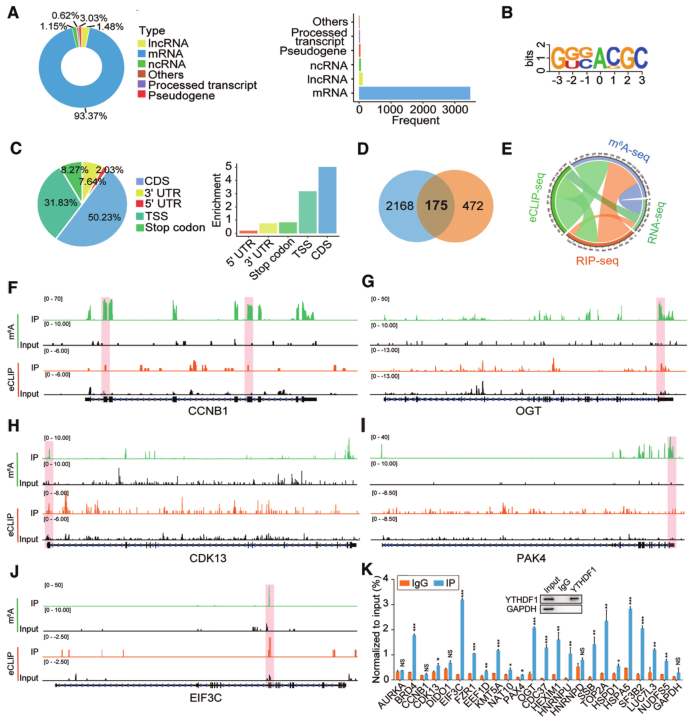
6.YTHDF1调节卵巢癌细胞EIF3C表达及整体翻译输出
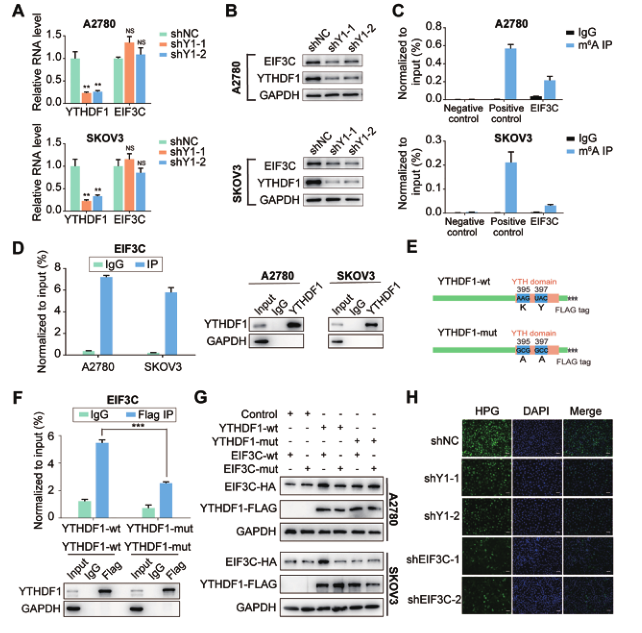
为了证实YTHDF1调节卵巢癌细胞中EIF3C的表达,我们首先检测了EIF3C在YTHDF1缺失时的转录和翻译。正如预期的那样,YTHDF1沉默降低了A2780和SKOV3卵巢癌细胞中EIF3C的蛋白质丰度,而不影响其RNA水平(图6A和B)。然后我们通过基因特异性的m6A分析来评估EIF3C mRNA的m6A修饰状态,观察到EIF3C mRNA显著富集(图6C)。此外,RIP-qPCR证实了A2780和SKOV3卵巢癌细胞中YTHDF1和EIF3C mRNA之间的相互作用(图6D)。
随着m6A修饰了EIF3C mRNA,我们接下来探究调节EIF3C表达的YTHDF1是否依赖于m6A。结果表明,卵巢癌细胞被YTHDF1宽型(YTHDF1 wt)或YTHDF1 mut结构体转染(图6E)。随后,用抗FLAG抗体和qPCR检测发现,在转染YTHDF1 wt的细胞中,EIF3C mRNA被有效地免疫沉淀,但是YTHDF1突变体和EIF3C mRNA之间的相互作用显著降低(图6F)。Western blot分析显示YTHDF1-wt能够增强EIF3C-wt的表达,而YTHDF1-wt对EIF3C-mut的表达有轻微的影响(图6G)。
由于EIF3C是蛋白质合成所必需的EIF3复合物的关键亚单位,我们接下来探讨YTHDF1是否能影响卵巢癌细胞的整体翻译输出。结果表明,与对照细胞相比,在EIF3C基因敲除后,合成蛋白质的HPG含量下降。类似地,与对照细胞相比,缺乏YTHDF1的细胞显示出HPG信号下降(图6H),表明YTHDF1可能影响整个蛋白质合成。总之,这些数据表明,YTHDF1通过调节卵巢癌细胞中EIF3C蛋白的表达促进整体蛋白合成。
7.EIF3C在卵巢癌细胞中起致癌作用
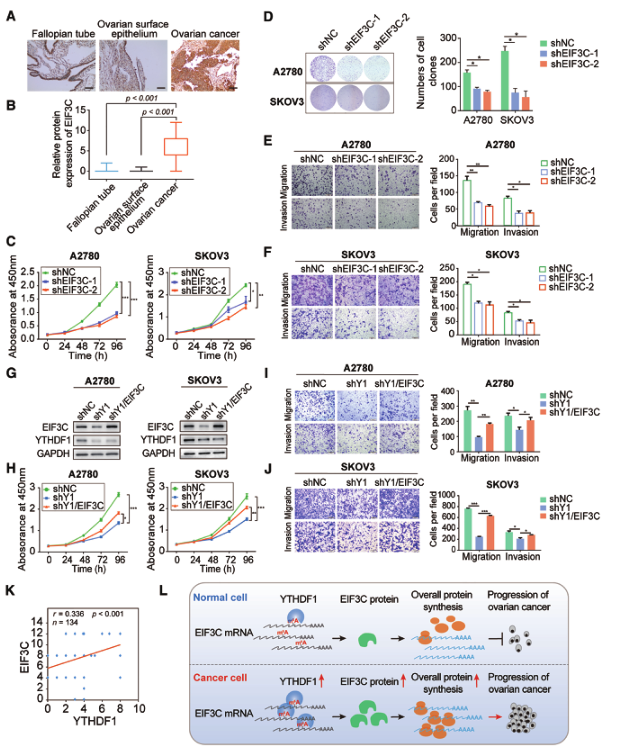
由于EIF3C在卵巢癌细胞中的作用尚不清楚,我们检测了EIF3C的表达水平。与输卵管相比,卵巢癌中EIF3C的蛋白表达显著升高(图7A和B)。为了进一步研究EIF3C在卵巢癌中的作用,我们分析了A2780和SKOV3细胞的细胞表型。两种卵巢癌细胞系中的细胞生长和集落形成能力在EIF3C基因敲除后均明显受到抑制(图7C和D)。此外,EIF3C缺乏也减少了卵巢癌细胞的迁移和侵袭(图7E和F)。这些结果表明EIF3C促进卵巢癌细胞的癌变。
接下来,我们在YTHDF1缺陷的A2780和SKOV3细胞中过表达了EIF3C,并且在两个细胞系中都恢复了EIF3C的表达(图7G)。YTHDF1基因敲除损伤细胞生长和菌落形成,而EIF3C的过表达可以逆转这种影响(图7H)。在EIF3C过表达后,由YTHDF1缺失抑制的细胞迁移和侵袭被重新建立(图7I和J)。这些结果提示EIF3C是YTHDF1促进卵巢癌进展的关键下游靶点。另外,YTHDF1的蛋白质丰度与EIF3C的表达呈正相关(图7K)。总之,增加YTHDF1的表达通过调节EIF3C的翻译增强了整体蛋白的合成,并促进了卵巢癌细胞的肿瘤发生(图7L)。
结论:
我们的研究发现,EIF3C是卵巢癌细胞中YTHDF1的直接靶点。YTHDF1以m6A依赖的方式控制EIF3C的翻译,并作为致癌调节因子影响整个蛋白质翻译。因此,靶向YTHDF1可能是卵巢癌治疗的一个有前途的候选方案。