






44分——ceRNA机制的正反馈环
大量研究表明circRNA和m6A甲基化修饰参与了包括肝细胞癌(HCC)在内的多种恶性肿瘤的病理进展和转移。但是circRNA如何和m6A甲基化修饰形成发聩环作用于HCC进展尚不清楚。本研究证实circGPR137B通过circGPR137B/miR-4739/FTO反馈环促进HCC的肿瘤发生和转移。这种通过circRNA和m6a修饰事件之间的功能耦合执行的正反馈机制为表观遗传学提供了一种新的模型机制。本研究于2022年6月发表在《Molecular Cancer》IF:41.444期刊上。
技术路线

主要实验结果:
1、circGPR137B下调和HCC患者的不良预后相关
对3对HCC肿瘤组织和配对癌旁组织进行测序,其差异表达circRNA热图如图1A所示,在P < 0.05和FC > 1.5的标准筛选后,在HCC中得到96个上调和51个下调的circRNA(图1B)。火山图显示hsa_circ_0017114,一个来源于线性RNA GPR137B的circRNA,在HCC中显著下调表达,将其命名为circGPR137B(图1C)。circRNA表达谱和RT-qPCR均表明相比于癌旁组织circGPR137B在HCC中下调(图1D-1E)。随后在87对HCC和癌旁组织中进一步证实了这个结果(图1F-1G)。预后分析发现和circGPR137B高表达组比较,circGPR137B低表达组具有较差的生存预后(图1H-1I)。以上结果表明circGPR137B下调和HCC患者的不良预后相关。
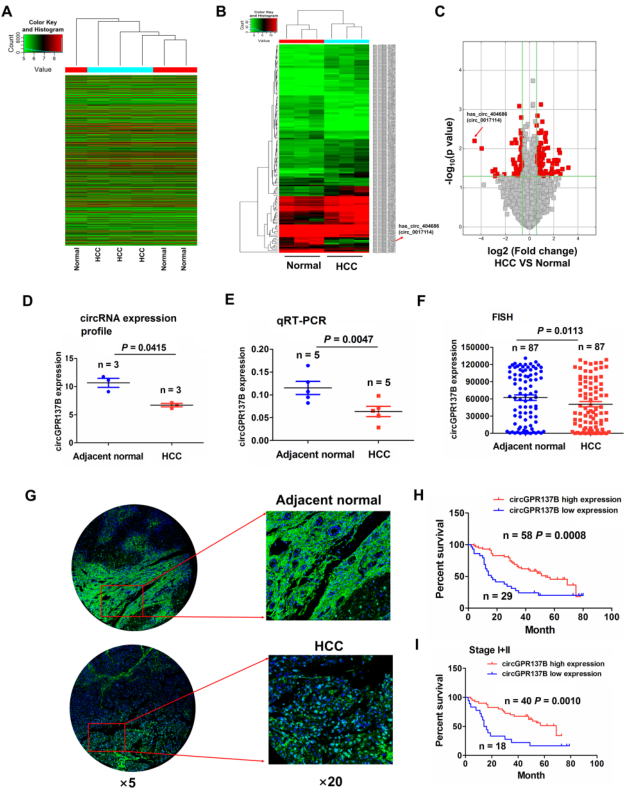
图1 circGPR137B下调和HCC患者的不良预后相关
hsa_circ_0017114来源于14q1(q42.3)染色体上GPR137B的外显子1,7区域,命名为circGPR137B,其基因组序列是40204nt,剪切长度是1537nt(图2A)。HepG2和Hep3B细胞在RNase R中暴露2小时,发现GPR137B的表达显著下降而circGPR137B的表达因其抗性而无明显改变(图2B)。随后探究了circGPR137B在actinomycin D暴露中的稳定性,结果显示circGPR137B的半衰期显著长于GPR137B(图2C)。RT-qPCR和FISH实验表明HCC组织中circGPR137B主要定位于染色质(图2D-2E)。
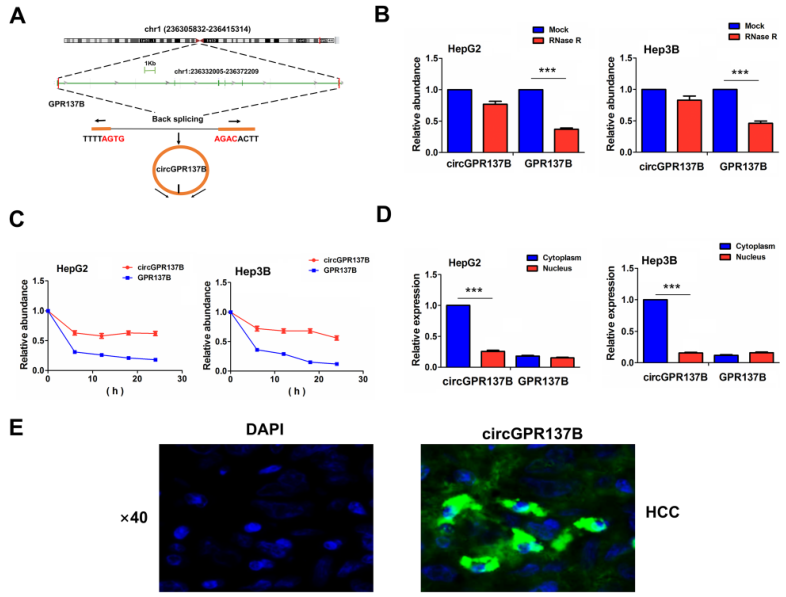
图2 HCC细胞中circGPR137B的特征
3、在体外circGPR137B抑制HCC细胞的生长和侵袭
作者检测了6株HCC细胞系中circGPR137B的表达,结果发现与正常肝细胞系比较,其在SK-hep-1细胞系中表达最高,在Hep3B中的表达最低(图3A)。所以后续在SK-hep-1细胞系中进行circGPR137B的敲低实验,在Hep3B中进行circGPR137B过表达实验(图3B)。MTT,集落形成和Transwell实验显示circGPR137B过表达HCC细胞的生长和侵袭,circGPR137B敲低则效果相反(图3C-3E)。以上表明circGPR137B在HCC中发挥抑癌作用。
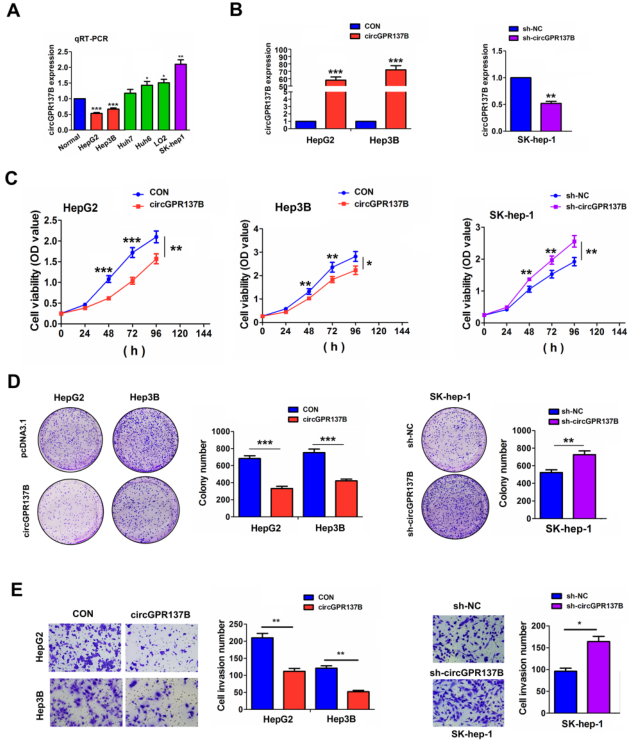
图3在体外CircGPR137B抑制细胞生长
4、在HCC中miR-4739的表达和circGPR137B负相关和HCC不良预后相关
基于circRNA表达谱和miRbase在线预测发现circGPR137B拥有5个潜在结合的miRNA(图4A),它们的结合位点如图4B所示。双荧光素酶实验结果显示circGPR137B的荧光素酶活性只在miR-4739 mimics处理时显著降低而非另外4个miRNA(图4C)。FISH结果显示相比于癌旁组织miR-4739的表达在HCC中显著上调(图4D-4E),并且miR-4739表达和circGPR137B显著负相关(图4F)。TCGA数据也证实miR-4739表达在HCC中显著上调,且和不良预后相关(图4G-4H)。以上说明HCC中miR-4739高表达且预示不良预后。
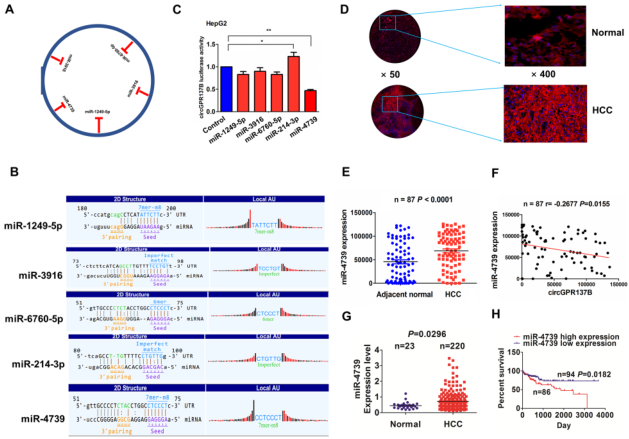
图4 在HCC中CircGPR137B与miR-4739呈负相关
5、在HCC中circGPR137B是miR-4739的海绵
基于上述结果作者选择miR-4739用于进一步实验。过表达circGPR137B显著降低miR-4739的表达(图5B)。此外,野生型circGPR137B和miR-4739的结合位点如图5A所示,同时构建了结合位点突变的circGPR137B荧光素酶报告基因载体。结果显示在野生型中,circGPR137B的荧光素酶活性在miR-4739过表达组显著降低,但是miR-4739突变后不改变,此外,在circGPR137B突变体中,miR-4739过表达不影响circGPR137B的荧光素酶活性(图5C)。AGO2-RIP显示和对照组比较内源性circGPR137B和miR-4739的富集可被Ago2抗体拉下(图5D)。FISH分析发现circGPR137B和miR-4739具有明显的细胞质共定位(图5E)。此外,细胞功能实验表明miR-4739过表达可显著抵消circGPR137B过表达对HepG2和Hep3B细胞增殖和侵袭的抑制效果(图5F-5G)。这些结果表明在HCC中circGPR137B可靶向调控miR-4739。
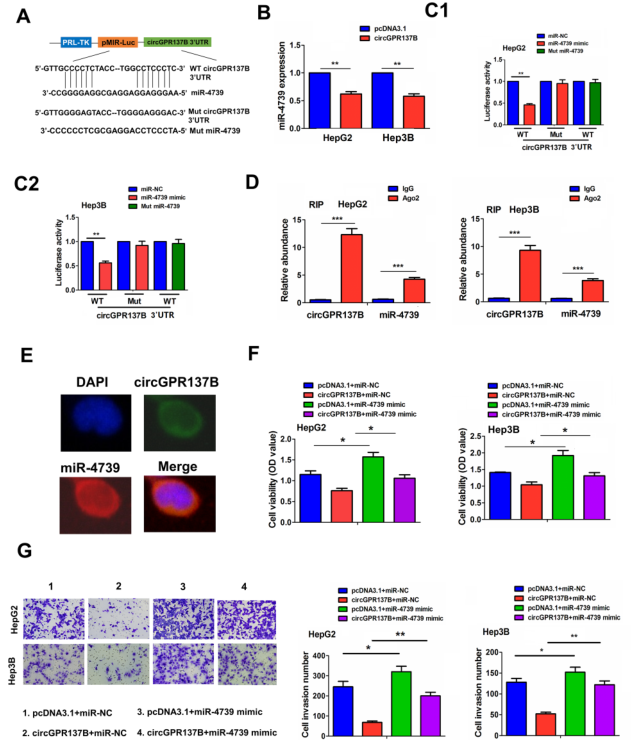
图5 在HCC中circGPR137B是miR-4739的海绵
6、在HCC中circGPR137B/miR-4739轴上调FTO
根据TargetScan7.1的结果,FTO可能是miR-4739的靶基因(图6A)。荧光素酶实验证实FTO和miR-4739可相互结合(图5B),并且miR-4739过表达可显著降低FTO,p21,p27,caspase-3/9和E-cadherin的表达,升高N-cadherin和vimentin的表达(图6C-6D)。此外,miR-4739过表达也可逆转circGPR137B对上述基因表达的影响。共转染FTO质粒和miR-4739 mimics后可观察到FTO过表达显著阻止了miR-4739过表达带来的促肿瘤作用(图6E-6G)。这些结果提示在HCC中circGPR137B/miR-4739轴上调FTO。
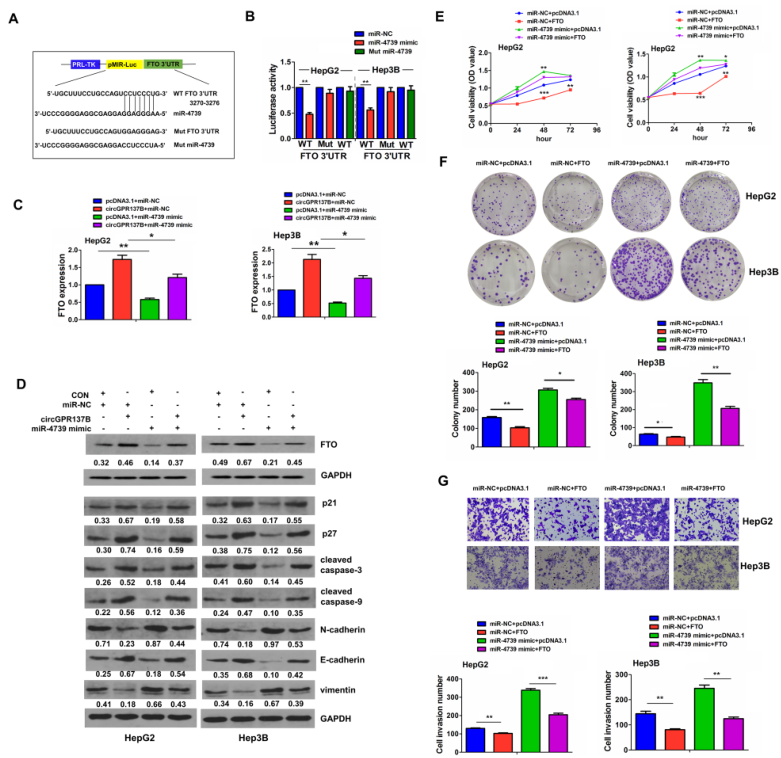
图6 在HCC中circGPR137B/miR-4739轴上调FTO
随后作者分析了FTO在HCC中的表达模式和预后价值,结果和预期一致,FTO表达在HCC组织中显著下降,且FTO下调与HCC预后不良有关(图7)。
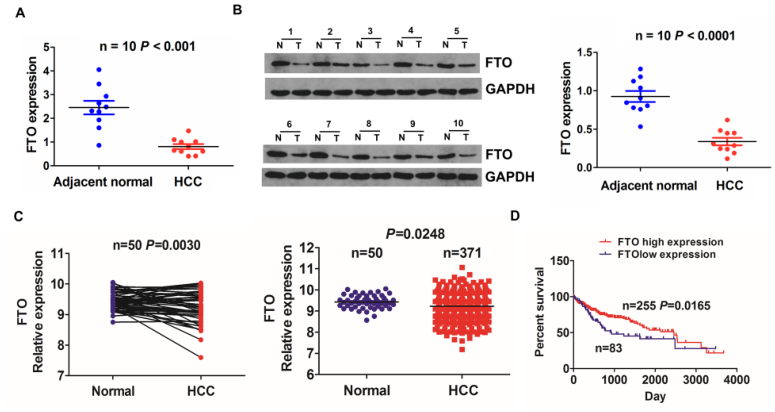
图7 FTO下调提示HCC预后不良
7、FTO介导circGPR137B的m6A修饰与circGPR137B形成正反馈环路
此前有研究表明肿瘤中circRNA会发生m6A修饰,而FTO是去甲基化酶,所以作者猜想FTO是通过介导circGPR137B的m6A修饰参与HCC进展。在HepG2和Hep3B细胞中,过表达FTO均可显著降低总体的m6A水平(图8A-8B)以及circGPR137B的m6A水平(图8C),但显著增加了circGPR137B的表达水平(图8D)。敲低FTO则降低circGPR137B的表达(图8E)。此外,敲低circGPR137B的表达也可显著增加miR-4739的表达增加FTO的表达(图8F),而miR-4739抑制剂处理则增加FTO的表达(图8G)。RIP实验揭示FTO可以结合circGPR137B(图8H)。重要的是,敲低circGPR137B可显著逆转FTO过表达引起的抗肿瘤效果(图8I-8J)。以上说明,FTO介导circGPR137B的m6A修饰与circGPR137B形成正反馈环路调控HCC进展。
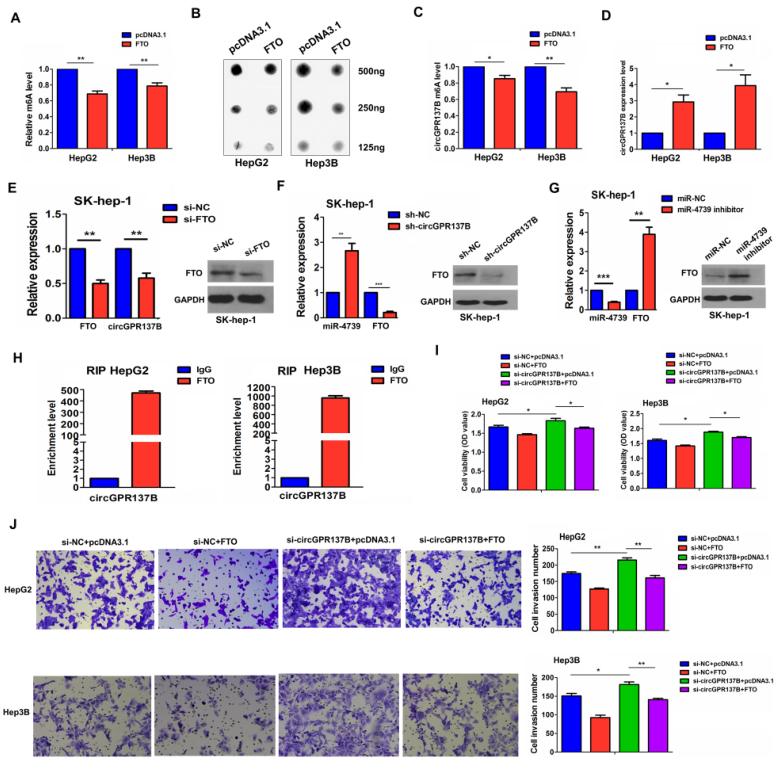
图8 FTO介导circGPR137B的m6A修饰与circGPR137B形成正反馈环路
如图9和图10所示,过表达circGPR137B显著增加HCC肿瘤的体积和重量,促进病理性改变,增加肿瘤增殖和FTO表达,同时也显著促进HCC细胞的肺转移结节形成,而敲低circGPR137B表达则效果完全相反。表明在体内circGPR137B抑制HCC肿瘤发生和转移。
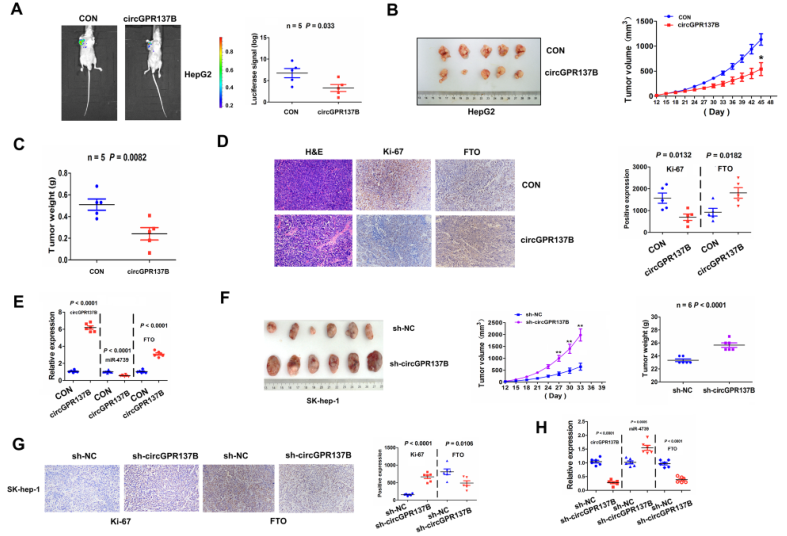

图10 CircGPR137B抑制体内HCC的肺转移
综上所述,本研究证实circGPR137B通过正反馈回路抑制肝癌的增殖和转移。CircGPR137B 通过海绵吸附 miR-4739 诱导 FTO表达。FTO蛋白进入细胞核介导circGPR137B的m6A去甲基化并促进其表达,因此,对circGPR137B 产生形成正反馈,从而强烈抑制肝癌细胞增殖、集落形成、侵袭和肺迁移。
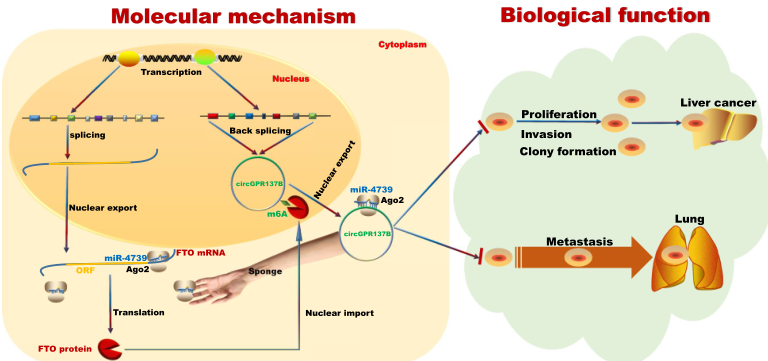
图11本研究的机制模拟图
参考文献:
Liu Lianyong., Gu Mingjun., Ma Junhua., Wang Ying., Li Miao., Wang Hui., Yin Xin., Li Xiangqi.(2022). CircGPR137B/miR-4739/FTO feedback loop suppresses tumorigenesis and metastasis of hepatocellular carcinoma. Mol Cancer, 21(1), 149. doi:10.1186/s12943-022-01619-4