






10分免疫新思路——B细胞分化命运的早期机制
抗体介导的免疫是由B细胞分化为包括浆母细胞、记忆细胞和生发中心(GC)细胞在内的多个细胞亚群引发的。B细胞的分化轨迹是由转录因子决定的,然而很少有专门决定早期B细胞命运的机制被描述。本研究中,作者报道了一种抑制浆母细胞遗传程序并促进 GC B细胞命运的转录后机制。本研究于2022年5月发表在《Cell Reports》IF:9.995期刊上。
技术路线:

主要实验结果:
作者希望研究的是在最初的抗原相遇和GC形成之前的这段时期支持B细胞命运决定的机制和基因程序。为了该目的,对免疫响应后GC形成前的腘窝淋巴结的单细胞悬液进行scRNA-seq检测。利用UMAP降维分析了6263个细胞,识别了6个簇,并根据它们的基因表达特征对它们进行了注释(Figures 1A, 1B)。其中一个簇的特征是Igkc和Ighd高表达,Iglc1的低水平表达,并且由于 NP 特异性细胞在B1-8 hi转基因小鼠中表达Igl,所以这个簇被定义为对抗原没有反应的原始B细胞(Figures 1B, 1C)。有两个簇显示了Fas、Cd86和GC转录因子Bcl6的高表达,提示这些细胞处于向GC B细胞分化的过程中(Figures 1B, 1C)。另一个簇表达激活标记Cd86和Fas以及高水平的趋化因子受体Ccr6,这是早期形成记忆细胞(eMBC)的典型特征。一个簇表达Cd86和Fas,但缺乏GC标记,因此被定义为pre-GC细胞。从纯化细胞群的RNA-seq数据集中提取的基因特征用于验证每个集群的细胞分配(Figures 1C)。正如预期的那样,细胞周期分析表明,原始细胞和eMBC大部分是非周期细胞,而一些pre-GC细胞和大部分早期GC B细胞是积极增殖的(Figures 1D)。利用假时间顺序将细胞沿可能的发育轨迹放置,可以预测pre-GC簇可能产生GC B细胞以及eMBC细胞亚群(Figures 1E)。因此,使用scRNA-seq能够识别pre-GC和具有形成成熟GC细胞能力的早期GC B细胞。
为了确定可能指导早期B细胞向GC B细胞定型的分子机制,对在pre-GC 簇中高表达的基因进行了GO富集分析,与原始和eMBC簇相比。在符合这些标准的 991个基因中,359 个基因被注释为编码RBP的基因(Figures 1F)。为了进一步表征早期进入GC B细胞期间RBP的表达,查询了所有注释为编码RBP 的基因 (GO: 0003723)。该分析证实,pre-GC细胞以及早期GC簇的细胞表现出最高的编码RBP基因的表达水平(Figures 1G)。这些结果表明RBP在早期B细胞分化和GC形成中发挥作用。
有趣的是,编码m6a结合蛋白的Ythdf2是符合富集标准的基因之一(在从原始细胞到pre-GC细胞的过渡期间,表达显著增加)(Figures 1H)。通过对Ythdf2表达作为伪时间函数的分析发现,它在发育轨迹的早期上调,从pre-GC细胞开始,在早期GC亚群中保持上调(Figures 1H, 1I)。Ythdf蛋白家族包含3个同源基因,然而,在整个B细胞系中,Ythdf1和Ythdf3的表达显著低于Ythdf2(Figure 1I)。与此相一致的是,利用qRT-PCR和多个引物对分选的B1-8 hi B细胞进行分析,证实Ythdf2在免疫反应早期被诱导,并在抗原特异性B细胞中占主导地位(Figure1J)。这些结果表明Ythdf2h是naive B细胞通过pre-GC状态向早期分化的GC细胞转变的潜在调节剂。
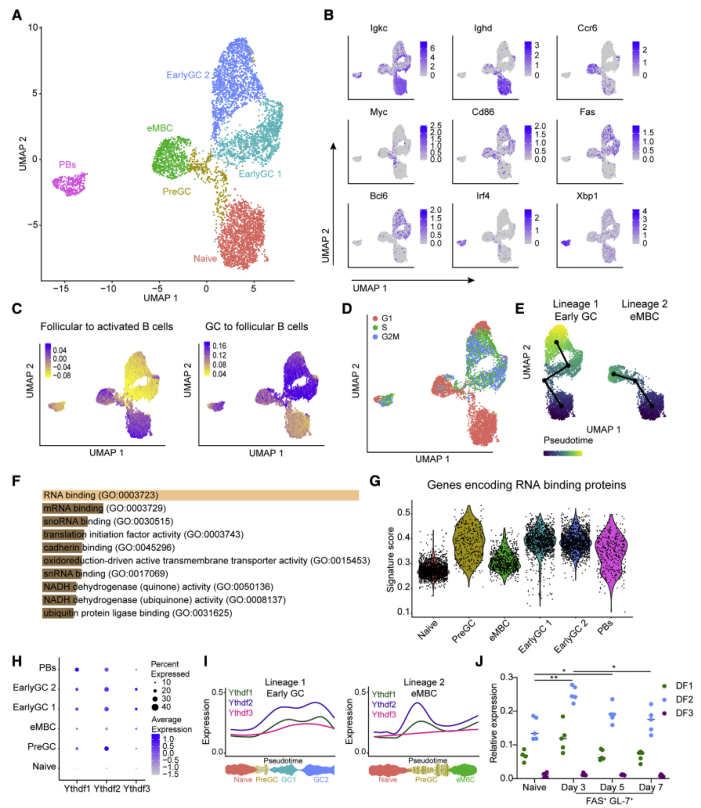
图1 YTHDF2在生发中心形成前B 细胞激活后上调表达
2、抗原特异性B细胞定位于淋巴结外滤泡区表达YTHDF2
接下来,确定YTHDF2蛋白在早期B细胞活化和GC形成过程中在B细胞中的表达模式。流式细胞术分析显示GL-7+ FAS+ B细胞在免疫应答第3天表达YTHDF2蛋白最多,第5天和第7天逐渐减少(Figure 2A)。在多克隆免疫反应中也观察到 YTHDF2 对疫苗接种的强烈上调,该免疫反应包括表达具有不同特异性和亲和力的免疫球蛋白的 B 细胞(Figure 2B)。因此,YTHDF2在抗原特异性B细胞免疫应答早期上调,并且在GC形成期持续表达。
为了检测B细胞在体内表达YTHDF2的部位,将表达YTHDF2-GFP的TdTomato+ B1-8hi B细胞转入宿主小鼠。在反应的第3天,约50%的TdTomato+ B1-8hi B细胞定位至靠近淋巴结包膜区域并高表达YTHDF2-GFP,并且其表达频率在淋巴结皮层深处的区域下降(Figures 2C, 2D)。运动B细胞的实时成像显示,YTHDF2存在于细胞的细胞质中,包括前缘和后缘;然而,由于尾缘含有较多的细胞质,大部分的YTHDF2蛋白质都位于这一区域(Figures 2E)。为了研究免疫球蛋白亲和性是否影响YTHDF2的表达,将B细胞转入表达低亲和性NP特异性BCRs (B1-8lo)和YTHDF2-GFP的小鼠,并将GFP表达与B1-8hi B细胞进行比较。结果发现,在NP-KLH免疫后3天,B1-8hi B细胞表达的YTHDF2水平高于低亲和力的细胞(Figure 2F)。综上所述,这些结果表明YTHDF2在位于淋巴结滤泡外区的B细胞中表达上调,且BCR亲和力调节其表达。
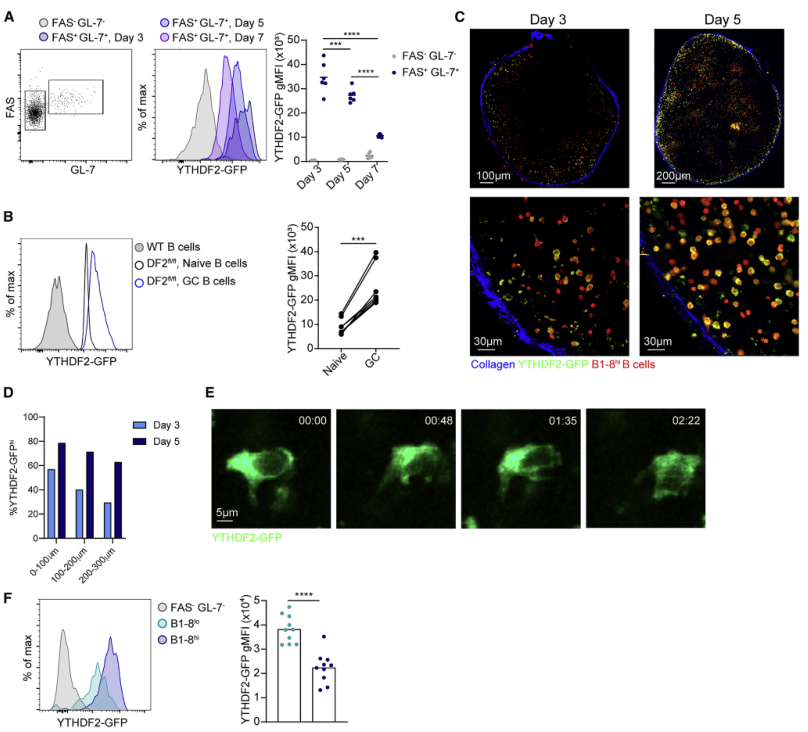
图2 YTHDF2蛋白表达在生发中心形成前上调
3、生发中心的形成依赖于YTHDF2
为了确定YTHDF2是否在抗体免疫反应中发挥作用,将Ythd2fl /fl小鼠与B细胞特异性CD23-Cre小鼠株(CD23-DF2fl/+和CD23-DF2fl/fl)杂交。ELISA结果显示,表达一个或两个Ythdf2-flox拷贝的小鼠血清IgM和IgG1水平较低,表明该基因是单倍不足的(Figure 3A)。为检测抗原特异性抗体的产生情况,用NP-KLH免疫小鼠,免疫1周和2周后ELISA检测血清中NP特异性抗体的存在情况。与野生型相比,CD23-DF2fl/fl小鼠在GC形成的第7天产生低水平的抗体;然而,这种差异在统计上并不显著(Figure 3B)。反应第14天GC形成后,CD23-DF2fl/fl中检测到极低水平的抗原特异性抗体,而CD23-DF2fl/+产生了结合总NPs和低密度NPs的IgG1(Figure 3B)。总的来说,这些实验表明,一个有效的抗体介导的免疫反应依赖于YTHDF2。
由于GC是抗体形成细胞的主要来源,接下来确定GC的形成是否依赖于YTHDF2。首先通过WT小鼠腹腔注射OVA启动,然后过继CD23-DF2fl/fl或WT B1-8hi naive B细胞,然后用NP-OVA增强(Figure 3C)。流式细胞术分析增强7天后腘淋巴结来源的细胞,结果显示,CD23-DF2fl/fl B1-8hi B细胞经过继后分化为GC细胞的比例比对照细胞低3.9倍(Figure 3D)。与对照组相比,缺乏Ythdf2的B细胞的PC/GC比率降低不明显,且PC/GC比率较高(Figure 3D)。流式细胞术分析少数CD23-DF2fl/fl B1-8hi GC B细胞未表达YTHDF2-GFP(Figure 3E)。为了进一步研究YTHDF2在GC形成中的意义,将WT和YTHDF2缺失的B1-8hi B细胞混合转移到WT宿主体内,然后给它们注射NP-OVA促进剂。7d后的流式细胞仪分析显示,在竞争条件下,Ythdf2缺陷的B1-8hi B细胞分化为GC细胞的能力完全受阻(Figure 3F)。因此,YTHDF2对产生完整的抗体介导的免疫反应至关重要,主要是通过促进GC的形成。
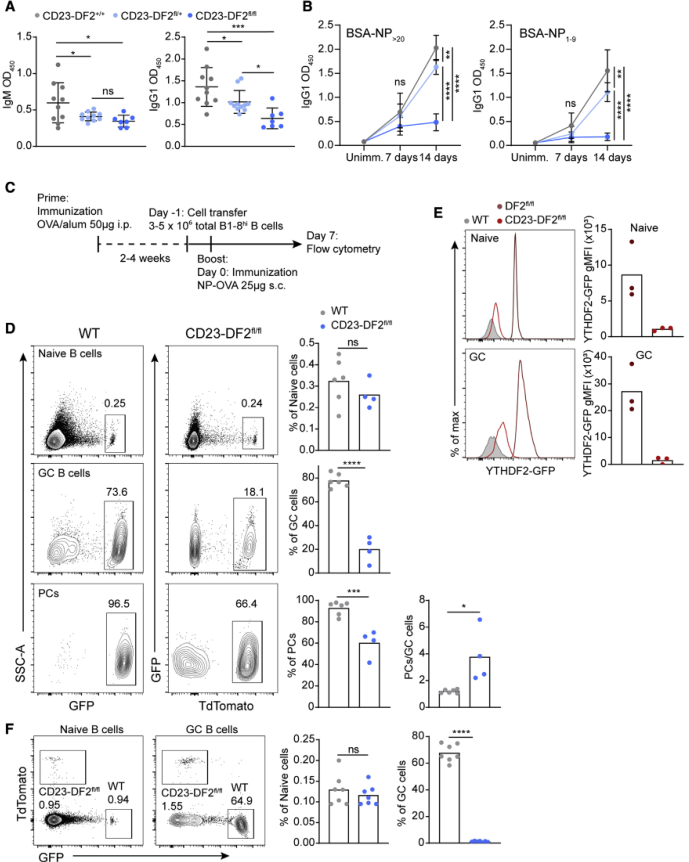
图3YTHDF2是有效的抗体介导的免疫反应和生发中心形成所必需的
4、YTHDF2是生发中心形成所必需的而不是早期B细胞激活所需
为了评估YTHDF2是否是早期阶段B细胞免疫应答所必须的,将WT和YTHDF2缺陷的B1-8hi B细胞一起转入宿主小鼠,并在NP-KLH免疫后3-7天通过流式细胞术检测FAS+ GL-7+ B细胞的存在。结果显示,在GC形成前的免疫反应第5天,YThdf2缺陷B细胞的频率显著降低(Figure 4A)。为了研究Ythdf2缺陷如何影响B细胞定位,使用TPLSM扫描完整的淋巴结。免疫后第5天淋巴结皮层可见CD23-DF2+/+ B1-8hi B细胞,第7天可见清晰的GC结构(Figure 4B)。CD23-DF2fl/fl B1-8hi B细胞转染小鼠后第5天检测到细胞簇;但2 d后未形成成熟的GC,提示GC形成需要YTHDF2(Figure 4B)。为了确定YTHDF2是在GC形成过程中发挥作用,而不是在同源抗原的初始激活过程中发挥作用,检测了YTHDF2缺陷的B细胞是否能够在体内对抗原作出反应。免疫16 h后,Ythdf2缺陷的B细胞检测到CD86、MHC-II、CD40等典型激活标志物表达上调(Figure 4C)。此外,CellTrace Violet追踪发现没有早期B1-8hi B细胞增殖缺陷(Figure 4D)。此外,在体外,通过增加anti-IgM剂量刺激B细胞,并没有发现任何显著的增殖缺陷或CD86表达的Ythdf2缺陷B细胞(Figure 4E)。 因此,BCR参与触发的B细胞活化和增殖不依赖于YTHDF2。
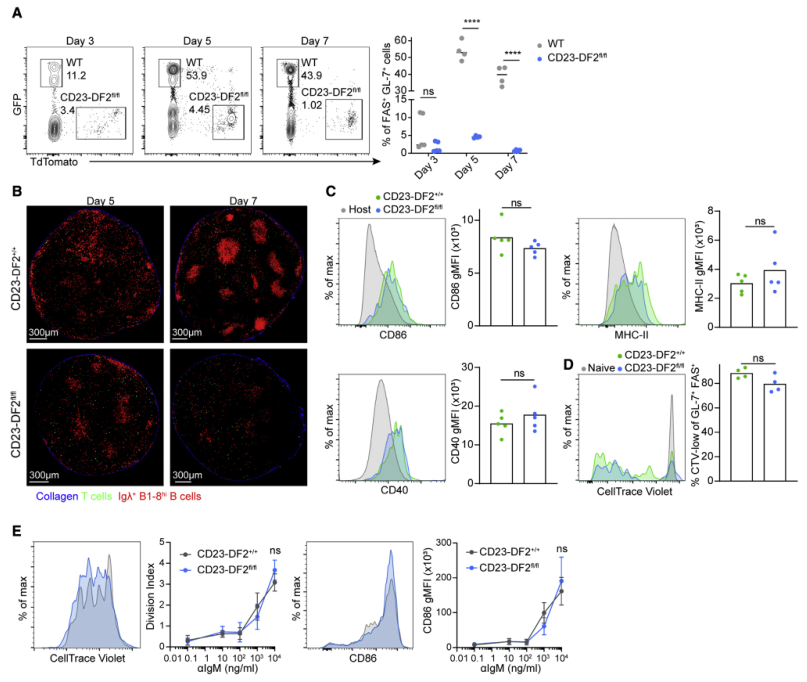
图4 YTHDF2是早期B细胞免疫响应所必需的而不是初始的B细胞激活所需
5、YTHDF2 抑制GC形成期间浆母细胞遗传程序
体内实验之间的差异表明,GC 形成对 YTHDF2 有很大依赖性,而在没有 YTHDF2 的情况下,早期 B 细胞活化和增殖没有受到干扰,这表明这种 m6A 阅读器在 B 细胞分化和命运决定中发挥作用,而不是在细胞周期进程和克隆扩增中发挥作用。为了进一步评估YTHDF2对B细胞发育哪个阶段至关重要,研究了支持B细胞命运的遗传程序。筛选WT和Ythdf2缺陷的B1-8hi B细胞,这些细胞在免疫接种5天后对受体宿主的抗原产生反应,并对它们进行RNA-seq。总计获得159个差异表达基因,值得注意的是,PB和PC中典型表达的基因,如Irf4、Xbp1、Sdc1和Prdm1,在Ythdf2缺陷的GL-7+ FAS+ B细胞中高表达(Figures 5A and 5B)。这些变化反映在GSEA揭示的ASC基因信号的表达增加(Figures 5C)。因此,YTHDF2在B细胞免疫反应的早期阶段抑制PB基因程序。
为了进一步研究在缺乏YTHDF2的情况下早期反应的B细胞的细胞命运,进行了scRNA-seq分析,共有11227个细胞显示了类似于图1中描述的集群(Figure 5D)。然而,尽管Ythdf2缺陷的B细胞分化为pre-GC和eMBC亚群,但它们形成的早期GC B细胞明显较少(Figure 5E)。与转录组结果一致,发现响应的Ythdf2缺陷B细胞形成了相对更多的PC,与对照B细胞比较(Figure 5F)。接下来,研究了早期FAS+ GL-7+ Ythdf2缺陷的B细胞是否除了ASC基因表达增加外,还具有PB相关功能。为了确保亚型特异性转录本的检测,使用分选的多克隆naive B细胞作为阴性对照,PC作为阳性对照。如预期的,如果有sIgM转录本的话,naive B细胞表现得非常低,而PC显示sIgM与mIgM转录本的比例非常高(Figure 5G)。这些结果为YTHDF2在pre-GC和GC早期抑制PB遗传程序中的作用提供了额外的证据。
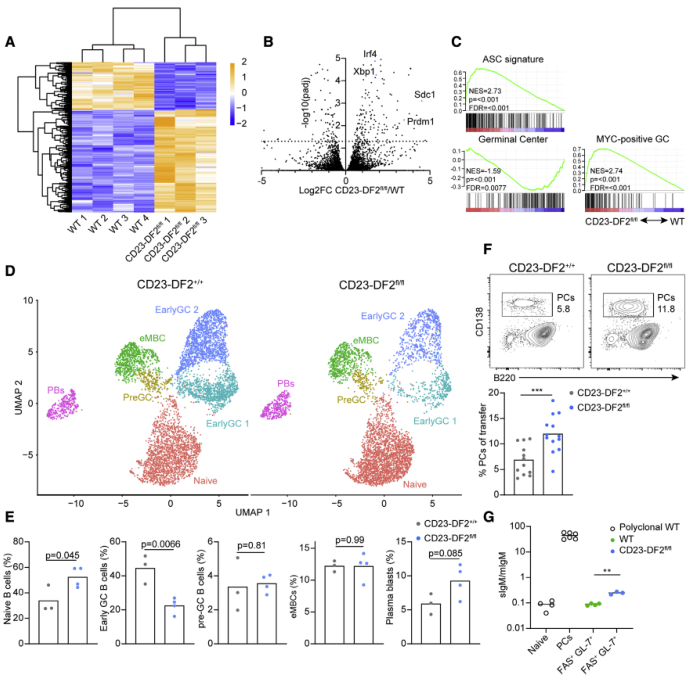
6、YTHDF2直接与浆母细胞调节基因的甲基化mRNA相互作用
由于前文的研究结果表明YTHDF2在抑制PB遗传程序中发挥作用,作者试图确定该蛋白是否可以直接靶向PB促进基因。为此,用脂多糖刺激分离的B细胞3天,在体外生成表达PB基因的细胞。然后,使用抗m6a抗体进行RNA免疫沉淀,并使用高通量测序作为读数。IP与对照组的比较显示,在Ythdf2缺陷B细胞中上调的几个PC分化基因,包括Prdm1、Sdc1、Xbp1和Irf4,显示出一个或多个m6A峰(Figure 6A)。此外,通过分析已发表的YTHDF2 IP在B细胞发育过程中的数据集发现Irf4和Xbp1转录本都直接与YTHDF2结合(Figure 6B)。Sdc1也与YTHDF2结合;然而,这个峰值在缺乏mRNA甲基化机制的细胞中持续存在,表明非特异性结合。从LPS刺激的转基因B细胞中提取HA-YTHDF2-GFP进行RNA免疫沉淀,然后进行qRT-PCR,得到类似的结果(Figure 6C)。这些发现表明YTHDF2有可能在转录后水平直接调节PC分化基因。
在早期反应的B细胞中上调的PB相关基因中,Irf4被认为是最上游的B细胞命运调控因子。免疫后5天对CD23-DF2+/+和CD23-DFfl/fl细胞中IRF4蛋白表达的分析表明,与对照相比,CD23-DFfl/fl 细胞中显示高水平 IRF4 的活化 CD138 阴性 B 细胞的频率是对照组的 2.5 倍,并且通过平均荧光强度测量检测到每个细胞的蛋白质水平增加(Figure 6D)。激活的和Ythdf2缺乏的B细胞中也检测到MYC蛋白水平的小幅升高(Figure 6E)。这些结果表明,YTHDF2具有通过调节IRF4 mRNA和蛋白表达水平直接调控PC形成的潜力。
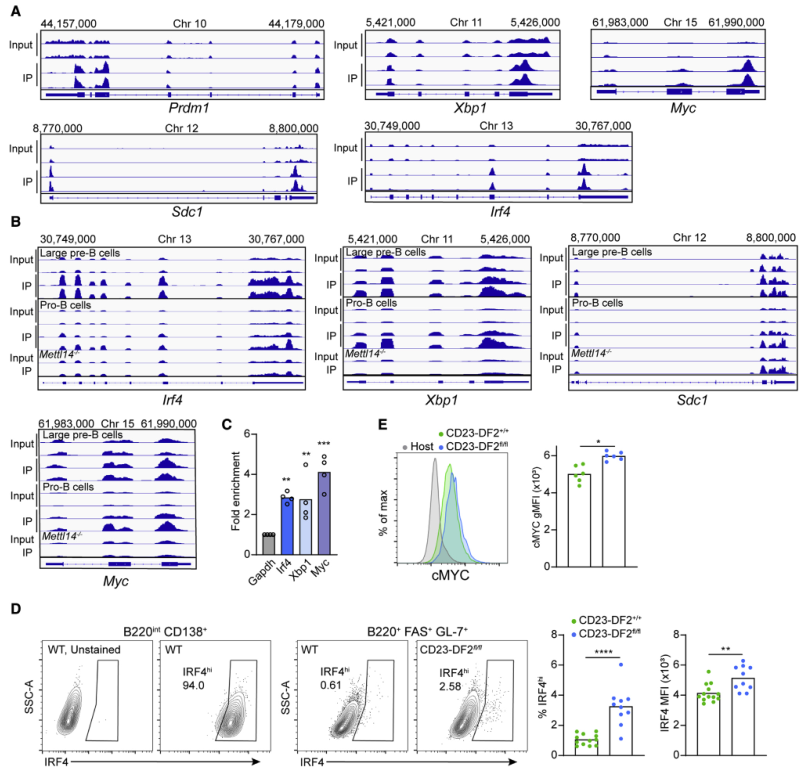
图6浆母细胞调节基因转录物是 YTHDF2 的直接靶标
7、生发中心和浆母细胞的形成不依赖于YTHDF1或YTHDF3
YTHDF蛋白在结构上非常相似,几项研究表明,所有三个同源段都促进RNA降解,并可以补偿任何同源段的损失。由于本文的结果显示Ythdf2的一个等位基因不足以产生完整的抗体介导的免疫反应,所以测试了YTHDF1和YTHDF3是否也有助于GC的形成。与Ythdf2缺陷B细胞相反,过继转染Ythdf1-和Ythdd3缺陷B1-8hi B细胞可有效生成GC细胞和PC细胞(Figures 7A and 7B)。在竞争条件下检测到Ythdf1缺陷的PCs和GC细胞频率较高,而Ythdf3缺陷的B细胞这些细胞亚群的频率没有变化(Figures 7C and 7D)。这些结果表明,与YTHDF2相比,Ythdd1/3序列不通过对B细胞中YTHDF总蛋白库的贡献来促进GC B细胞的形成。流式细胞仪检测免疫后第3、5、7天B1-8hi B细胞,显示FAS+ GL-7+ B1-8hi B细胞的形成不需要YTHDF1和YTHDF3(Figure 7E)。然而,来自免疫小鼠的FAS+ GL-7+ B1-8hi B细胞的RNA-seq实验显示,在Ythdf2缺陷B细胞中显著差异表达的基因在缺少Ythdf1和Ythdf3的情况下也发生了类似的改变,尽管程度较轻(Figures 7F and 7G)。虽然没有在Ythdf1和Ythdf3缺陷B细胞的基因表达数据集中检测到IRF4水平的增加,但在Ythdf3缺陷B细胞中观察到ASC信号的增加(Figures 7G)。因此,YTHDF2在B细胞中起主导作用,而非YTHDF1和YTHDF3,这是由于它在B细胞中高表达。
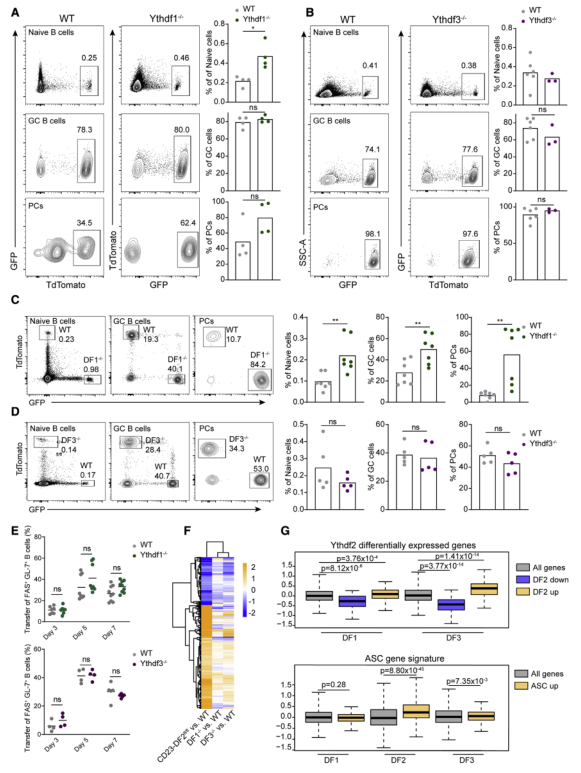
图7生发中心的形成不依赖于YTHDF1和YTHDF3
总之,本研究使用scRNA-seq 和转基因小鼠证明YTHDF2由早期反应的抗原特异性B细胞表达,并通过抑制与浆细胞形成相关的基因促进其分化为生发中心 B细胞,而其他YTHDF旁系同源物对于体液免疫反应是可有可无的。
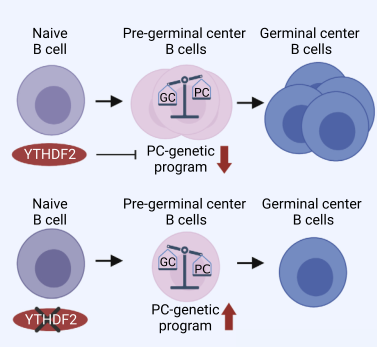
图8图文摘要
参考文献:
Grenov Amalie., Hezroni Hadas., Lasman Lior., Hanna Jacob H., Shulman Ziv. (2022). YTHDF2 suppresses the plasmablast genetic program and promotes germinal center formation. Cell Rep, 39(5), 110778. doi:10.1016/j.celrep.2022.110778