






m6A阅读器YTHDF3介导的PRDX3翻译减轻肝纤维化
PRDX3是线粒体氧化应激的主要调节因子,并发挥肝脏保护作用,但PRDX 3在肝纤维化中的作用尚不清楚。本研究旨在阐明PRDX3对肝纤维化的影响以及m6A修饰调节PRDX3的潜在机制。在动物模型和患者临床标本中,发现PRDX3表达与肝纤维化呈负相关。PRDX3沉默加剧了肝纤维化和肝星状细胞(HSC)激活,而HSC特异性过表达PRDX3减轻了肝纤维化。在机制上,PRDX3至少部分通过ROS/TGF-β1/Smad2/3途径抑制HSC激活。此外,PRDX3 mRNA被m6A修饰,并与m6A阅读器YTHDF1-3互作。更重要的是,当YTHDF3(而不是YTHDF1/2)被敲除时,PRDX3表达被抑制。PRDX3翻译以m6A依赖的方式直接受YTHDF3调节,从而影响其在肝纤维化中的功能。总的来说,PRDX3是肝纤维化的关键调节因子,在HSC中靶向YTHDF3/PRDX3轴可能是一种很有希望的治疗肝纤维化的方法。本文于2022年6月发表在“Redox Biology”杂志(IF= 10.787)上。
技术路线

结果
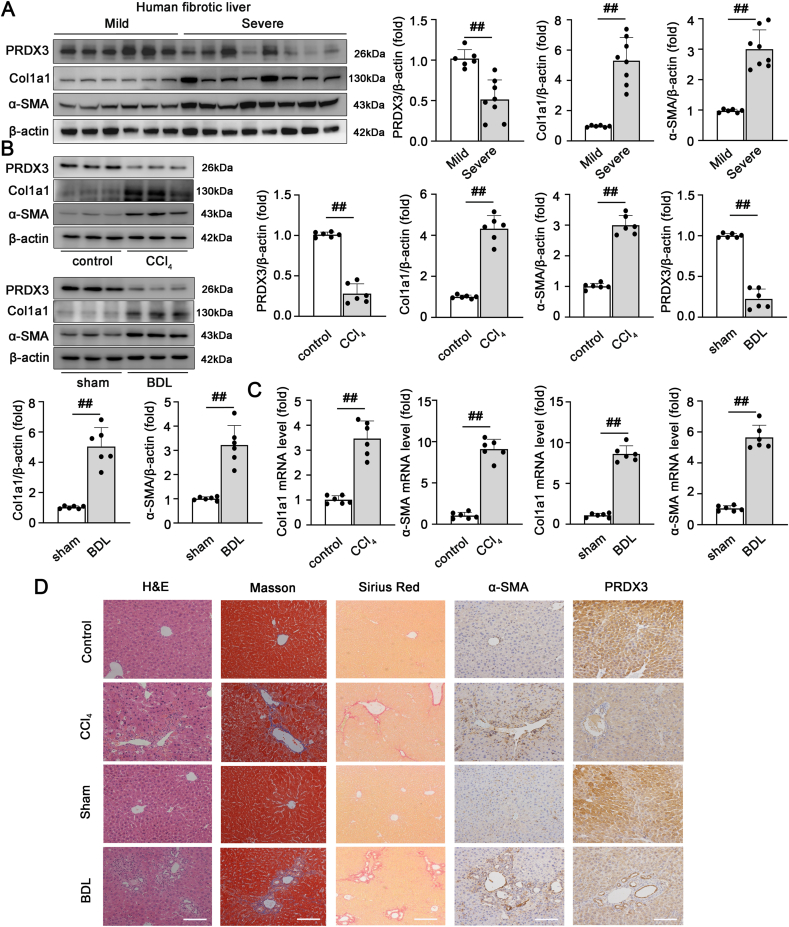
1)PRDX3在人和小鼠的纤维化肝组织中表达降低
我们最初检测了慢性肝纤维化患者肝组织中PRDX3的表达,以探讨PRDX3在临床肝纤维化中的潜在作用。重度纤维化患者的PRDX3蛋白水平明显低于轻度纤维化患者。在纤维化肝脏中检测到胶原和α-SMA的积聚(图1A)。这些结果表明PRDX3下调参与了人肝纤维化。然后,我们使用纤维化动物模型进一步确定PRDX3表达与肝纤维化之间的相关性。用CCl4处理的小鼠表现出显著降低的PRDX3蛋白和mRNA水平,这些降低伴随着纤维化标记物Col1a1和α-SMA的表达增加(图1B和C)。免疫组化(IHC)染色显示,CCl4诱导的小鼠PRDX3表达下降与纤维化肝脏中α-SMA表达呈负相关(图1D)。这些数据表明,PRDX3的表达与肝纤维化的严重程度呈负相关。
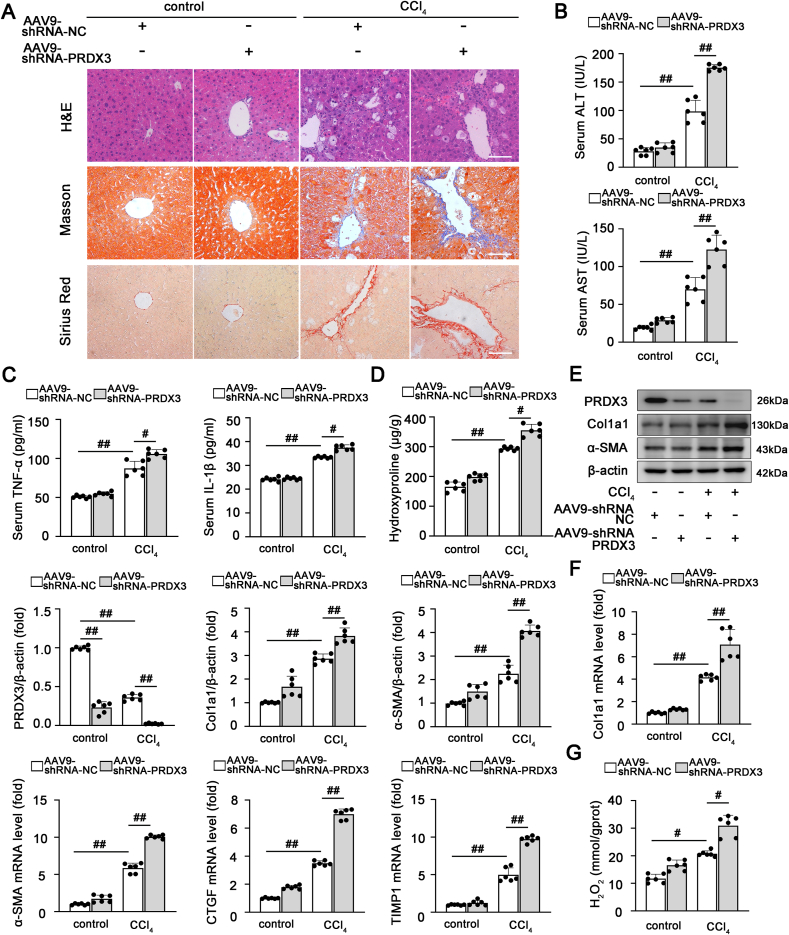
2)PRDX3下调加重肝纤维化
为了进一步了解PRDX3在肝纤维化中的作用,通过尾静脉将PRDX3-shRNA注射到小鼠体内。如图2A-C所示,PRDX3敲除加重了肝损伤,并增加了AST、ALT、TNF-α和IL-1β的水平。与这些发现一致,在注射PRDX3 shRNA的小鼠中检测到显著的纤维化诱导,表现为更强的Sirius Red and Masson染色,更高的羟脯氨酸含量,以及纤维化和HSC激活相关基因(Col1a1,α-SMA,CTGF和TIMP1)的表达水平增加(图2A,D-F)。此外,PRDX3敲除导致H2O2含量显著增加,这表明PRDX3沉默在肝纤维化期间诱导细胞内ROS积累(图2G)。基于这些结果,PRDX3敲除加重HSC激活和体内肝纤维化。
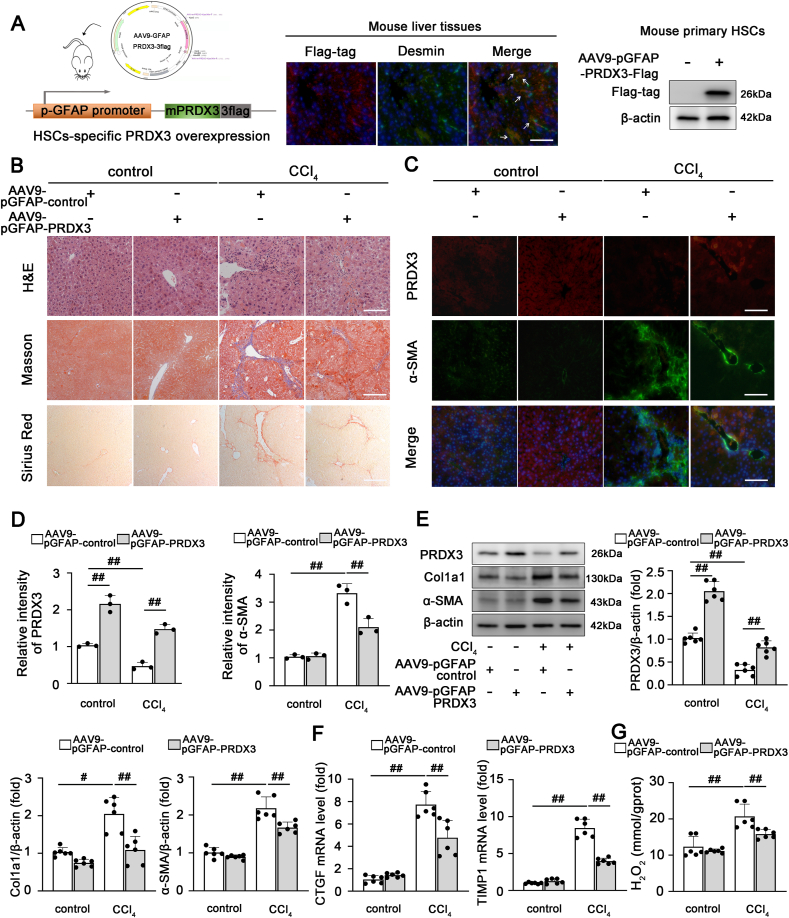
3)HSC特异性过表达PRDX3减轻肝纤维化
由于HSC激活是肝纤维化的关键因素,我们假设针对HSC中PRDX3的治疗可能预防肝纤维化。如图3A所示,外源性flag标记的PRDX3成功转染到HSC中。正如所料,PRDX3在HSC中特异性过表达显著减轻了肝损伤和纤维化,如肝脏HE的改善、Masson、Sirius Red和α-SMA-PRDX3染色的改善以及纤维化和HSC激活相关标记物的水平下降(图3B-F)。此外,在AAV9-pGFAP-PRDX3处理的小鼠中观察到H2O2含量降低,这表明PRDX3过表达减少了肝纤维化中的细胞内ROS积累(图3G)。因此,HSC特异性PRDX3过表达显著减轻小鼠肝纤维化。
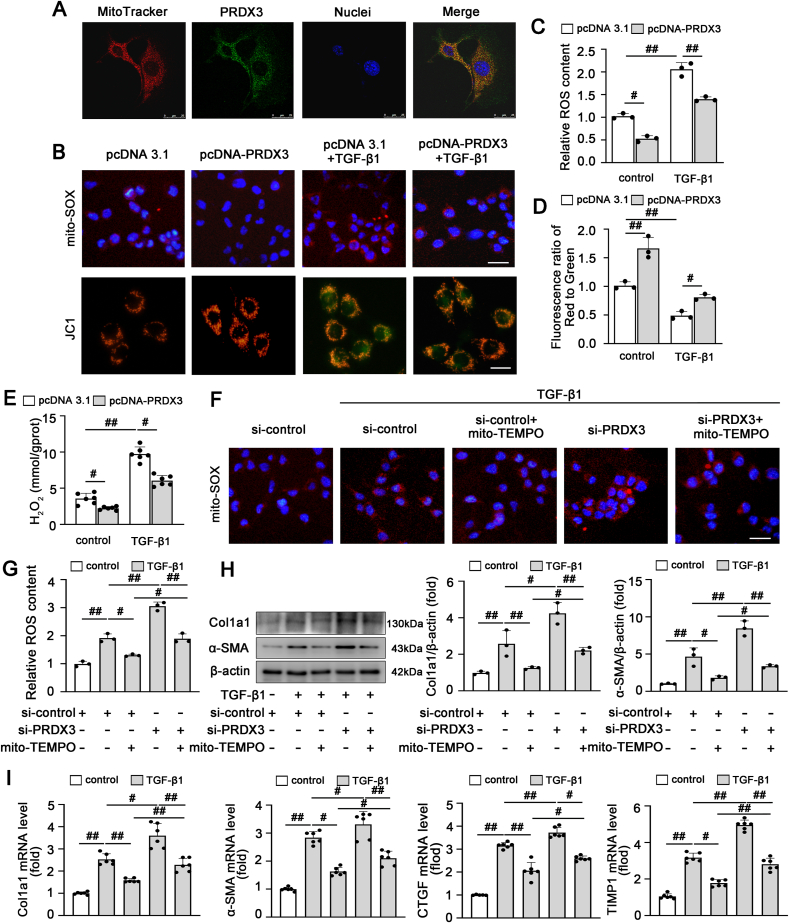
4)PRDX3通过调节线粒体ROS/TGF-β1/Smad2/3途径抑制HSC激活
我们进一步研究了PRDX3介导的线粒体ROS是否参与HSC激活。如图4A所示,PRDX3主要定位于LX-2细胞的线粒体中。经TGF-β1处理后,线粒体ROS水平、线粒体膜电位和H2O2含量增加,PRDX3过表达后显著降低(图4B-E)。PRDX3沉默导致活性氧生成旺盛,但在mito-TEMPO处理后,ROS生成减弱(图4F和G)。此外,mito-TEMPO显著减弱了PRDX3敲除引起的肝纤维化恶化和HSC激活(图4H和I)。这些结果表明,PRDX3减少肝纤维化中HSC激活的潜在机制可能主要通过抑制线粒体ROS的产生来介导。
越来越多的证据表明,线粒体ROS参与肝纤维化形成的机制是通过TGF-β1/ Smad2/3通路介导的。然后,我们探索了PRDX3介导的线粒体ROS生成调控是否与HSC中TGF-β1/Smad2/3通路相关。结果表明,HSC特异性过表达PRDX3可显著降低p-Smad2和p-Smad3的体内外表达水平。此外,PRDX3敲除增加了HSC活化相关基因和纤维化基因的水平,而TGF-β1抑制剂SB431542处理显著降低了这些增加(补充图,未展示在文章中)。这些数据表明,PRDX3通过抑制TGF-β1/Smad2/3通路减轻HSC活化和肝纤维化。
5)m6A修饰调节PRDX3的表达并影响其在肝纤维化中的功能
由于PRDX3是肝纤维化的关键调节因子,我们进一步探索了PRDX3上游的潜在机制。SRAMP预测PRDX3 mRNA中存在多个潜在的m6A位点;因此,我们推测PRDX3在肝纤维化中的调节依赖于m6A修饰。首先,我们检测了肝纤维化中m6A的水平。如图5A所示,在肝纤维化中观察到总RNA群体中m6A水平升高。随后,带有m6A特异性抗体的MeRIP qRT–PCR证明PRDX3 mRNA在LX-2细胞中显著富集(图5B),这表明PRDX3对肝纤维化的作用可能与其m6A修饰有关。此外,为了进一步研究m6A基序在PRDX3调节中的功能,用腺嘌呤-鸟嘌呤(A-G)突变替换了SRAMP预测的PRDX3中的三个m6A位点,如下所示:PRDX3-mut1(A399G)、PRDX3-mut2(A534G)、PRDX3-mut3(A1037G)和PRDX3-mut1-3(A399G、A534G、A1037G)。将这些突变体转染到LX-2细胞中。PRDX3突变体中的m6A水平明显低于野生型PRDX3(PRDX3-WT)(图5C和D)。更重要的是,PRDX3 mut1-3的m6A水平的下降程度大于只有一个潜在m6A突变的mRNA的下降程度,这表明三个m6A位点的合作是PRDX3 m6A修饰的关键机制(图5C)。此外,m6A基序突变后,PRDX3蛋白水平显著降低(图5E-G),这表明PRDX3的m6A修饰可能调节肝纤维化中的PRDX3蛋白质表达。然后,我们探讨了PRDX3的m6A修饰是否进一步调节了其对肝纤维化的保护作用。与使用PRDX3-WT获得的结果相比,PRDX3 m6A位点突变显著诱导HSC激活和肝纤维化(图5E-H)。基于这些数据,m6A修饰可能参与PRDX3的转录后调节,并改变其在肝纤维化中的表达和功能。
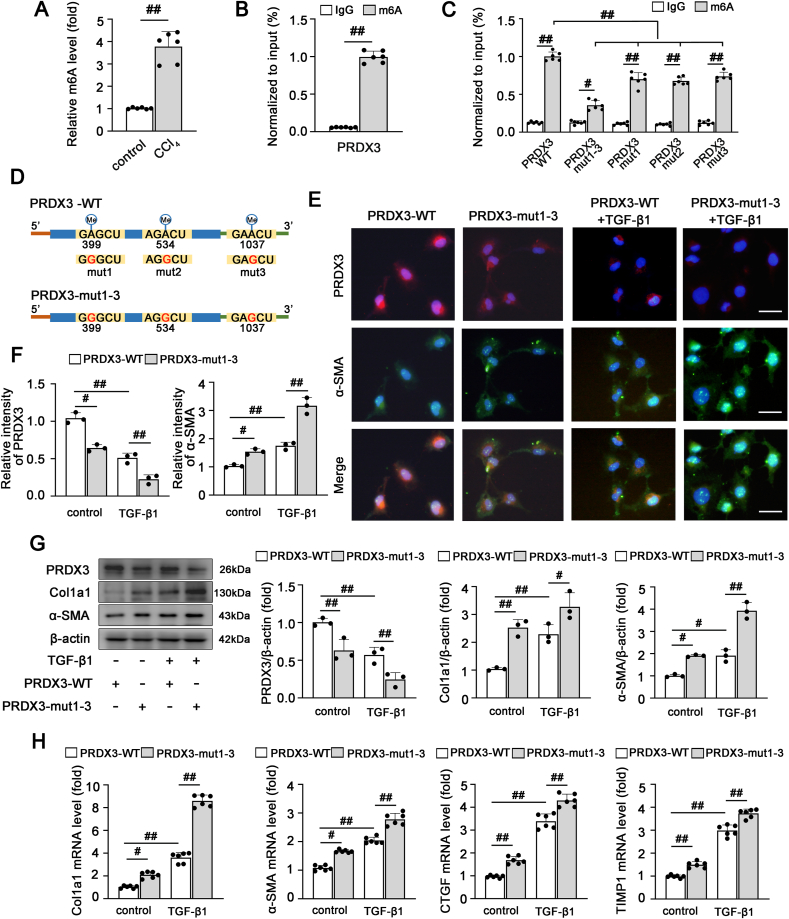
6)PRDX3 mRNA翻译由YTHDF3以m6A依赖的方式调节
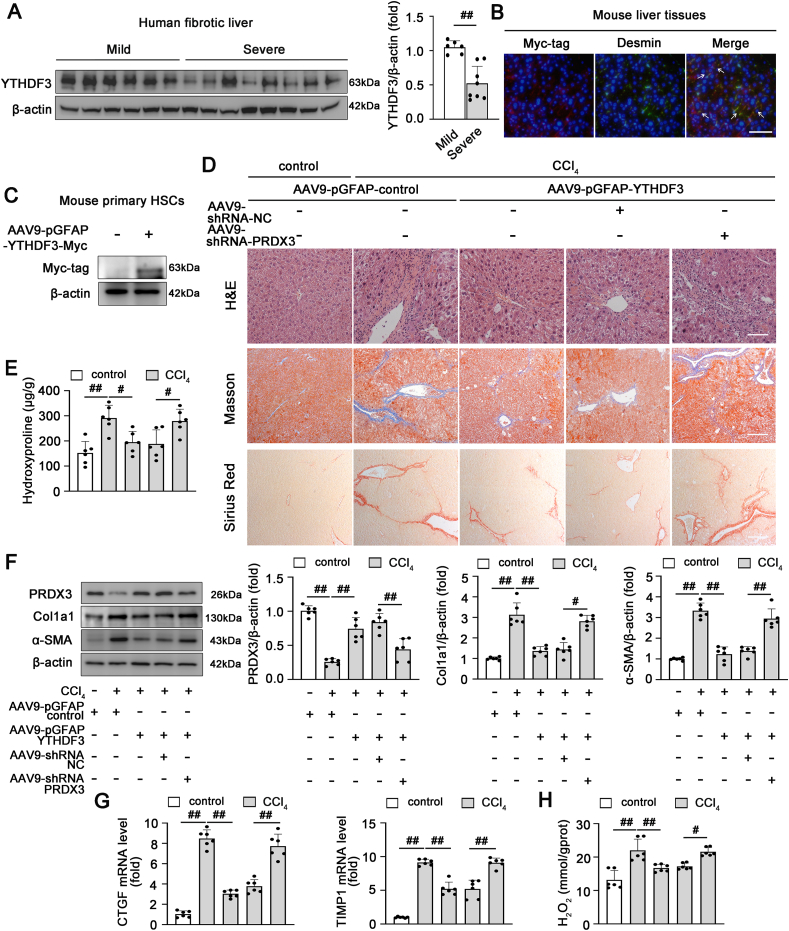
我们试图确定通过m6A修饰在肝纤维化中调节PRDX3表达的关键m6A阅读器。RNA下拉结合质谱法鉴定LX-2细胞中的PRDX3 mRNA相互作用蛋白(图6A)。在质谱结果中,YTHDF1-3和IGF2BP2-3特异性结合PRDX3 mRNA(图6B)。在肝纤维化过程中,YTHDF1-3表达明显下调,而IGF2BP2-3的表达没有显著差异,表明YTHDF 1-3可能是肝纤维化期间影响PRDX3 mRNA的潜在m6A阅读器(补充图,未展示)。此外,敲除YTHDF3而非YTHDF1/2抑制了PRDX3蛋白的表达(图6C)。相反,YTHDF3过表达显著增加了PRDX3蛋白的表达(图6D)。通过使用生物素化的PRDX3 mRNA进行YTHDF3下拉,进一步确定了YTHDF 3和PRDX3之间的特定相互作用(图6E)。RIP分析表明,与IgG免疫沉淀物相比,PRDX3 mRNA在含YTHDF3的免疫沉淀物中富集(图6F)。这些数据表明,YTHDF3与PRDX3特异性结合,并调节其在LX-2细胞中的表达。
由于YTHDF3通过其在YTH结构域中的m6A结合点(W438和W492)与m6A位点结合,W438与W492突变消除了YTHDF 3的mRNA结合能力。因此,用表达YTHDF3-WT或具有W438A和W492A突变的YTHDF3-mut的质粒转染LX-2细胞。RIP分析表明,在转染YTHDF3-WT的细胞中,PRDX3 mRNA有效免疫沉淀,而YTHDF 3-mut和PRDX3 mRNA之间的相互作用显著减少(图6G)。此外,YTHDF3-WT而非YTHDF 3-mut增加了PRDX3的表达。具有m6A位点突变的PRDX3在存在YTHDF3-WT过表达的情况下没有表现出显著的蛋白质水平增加(图6H),这表明YTHDF3介导的PRDX3表达调节依赖于m6A。由于YTHDF3增加了m6A修饰的mRNAs的翻译效率,我们随后试图确定YTHDF 3是否调节PRDX3的翻译。事实上,YTHDF3敲除抑制了PRDX3的翻译(图6I)。这些结果表明,在肝纤维化中,PRDX3 mRNA的翻译可能以m6A依赖性的方式受YTHDF3的调节。

7)PRDX3敲除抑制HSC特异性过表达YTHDF3对小鼠肝纤维化的保护作用
我们观察到人类和小鼠纤维化肝脏中的YTHDF3蛋白水平显著降低,这表明YTHDF3表达与肝纤维化的进展呈负相关(图7A)。此外,我们进一步探讨了YTHDF3是否通过调节PRDX3表达在肝纤维化中发挥作用。我们首先验证了HSC中YTHDF3过表达的特异性(图7B和C)。HSC特异性过表达YTHDF3显著减轻了肝损伤和纤维化(图7D-G)。值得注意的是,HSC中的YTHDF3过表达显著增加了PRDX3蛋白水平,并减少了细胞氧化损伤(图7F,H)。然而,PRDX3敲除可消除AAV9-pGFAP-YTHDF3对肝纤维化的保护作用(图7D-H)。这些数据表明HSC特异性过表达YTHDF3主要通过上调PRDX3表达显著减轻CCl4诱导的肝纤维化。
结论:PRDX3的HSC特异性过表达通过抑制线粒体ROS/TGF-β1/Smad2/3信号传导抑制HSC激活并改善肝纤维化。机制上,YTHDF3特异性调节PRDX3的翻译和表达,并在肝纤维化过程中影响其功能。我们的结果表明靶向HSC中的YTHDF3/PRDX3轴可能是一种有希望的治疗肝纤维化的策略。
参考文献:Sun R, Tian X, Li Y, Zhao Y, Wang Z, Hu Y, Zhang L, Wang Y, Gao D, Zheng S, Yao J. The m6A reader YTHDF3-mediated PRDX3 translation alleviates liver fibrosis. Redox Biol. 2022 Aug;54:102378. doi: 10.1016/j.redox.2022.102378.