






ALKBH3依赖的Aurora A mRNA的m1A去甲基化抑制纤毛生成
初级纤毛是触角状的亚细胞结构,作为信号平台,调节许多细胞过程和胚胎发育。m1A RNA修饰在RNA代谢和基因表达中起关键作用;然而,m1A修饰的生理功能在很大程度上仍不清楚。我们发现m1A去甲基化酶ALKBH3通过其去甲基化活性显著抑制哺乳动物细胞纤毛发生。机制上,ALKBH3去除纤毛发生的主抑制因子Aurora A mRNA上的m1A位点。ALKBH3的缺失增强了Aurora A mRNA的衰减并抑制其翻译。此外,在斑马鱼胚胎中,ALKBH3突变体表现出纤毛缺陷,包括弯曲体、心包水肿、异常耳石和前肾管扩张,野生型ALKBH3显著拯救了这些缺陷,但其催化失活突变体却不能。在脊椎动物细胞和胚胎中,由ALKBH3耗尽引起的纤毛缺陷也被Aurora A mRNA的异位表达显著逆转。总之,我们的数据表明,依赖于ALKBH3的m1A去甲基化在Aurora A mRNA的调控中起着至关重要的作用,这对脊椎动物纤毛发生和纤毛相关的发育事件至关重要。本文于2022年3月发表于Cell Discovery (IF=38.079)上。
技术路线:

主要研究结果:
(1) ALKBH3通过依赖m1A的方式抑制纤毛发生
为了研究m1A RNA修饰在纤毛发生中的作用,我们首先在正常条件下使用小干扰RNA (siRNA)对人类RPE-1细胞中编码m1A调节蛋白的靶向基因进行了功能筛选。TRMT6、TRMT61A或TRMT61B的缺失对RPE-1细胞纤毛发生没有显著影响(补充图未展示)。用两种siRNA中的一种沉默ALKBH1或TRMT10C对纤毛比例有影响。重要的是,用两种不同的siRNA敲除ALKBH3显著增加了纤毛细胞的百分比,但不影响纤毛的长度(图1a-d)。此外,ALKBH3-FLAG蛋白的异位表达显著逆转了ALKBH3敲除细胞中增强纤毛的作用(图1e-g)。为了确认ALKBH3在纤毛中的功能,我们外源表达了ALKBH3-FLAG,发现强制表达ALKBH3- FLAG显著减少了血清饥饿处理的RPE-1细胞的纤毛发生(图1h-j)。
由于ALKBH3在mRNA上的去甲基化活性被D193A的突变所抑制,我们制备了一个催化不活跃的ALKBH3突变体(ALKBH3D193A),以研究ALKBH3在纤毛发生中的调控是否依赖于它的去甲基化活性。通过LC-MS/MS对RPE-1细胞中m1A mRNA修饰的定量分析,证实我们构建的ALKBH3-D193A结构确实是催化活性不高的(补充图未展示)。野生型ALKBH3-FLAG的异位表达显著恢复了ALKBH3耗尽细胞的纤毛表型,但在RPE-1细胞中,突变型ALKBH3-D193A-FLAG的异位表达无效(图1k-m)。此外,过表达的ALKBH3-D193A突变体对血清缺失的RPE-1细胞的纤毛没有显著影响(补充图未展示)。总之,这些数据表明m1A RNA去甲基化酶ALKBH3在哺乳动物细胞中起着纤毛发生的负调控作用。
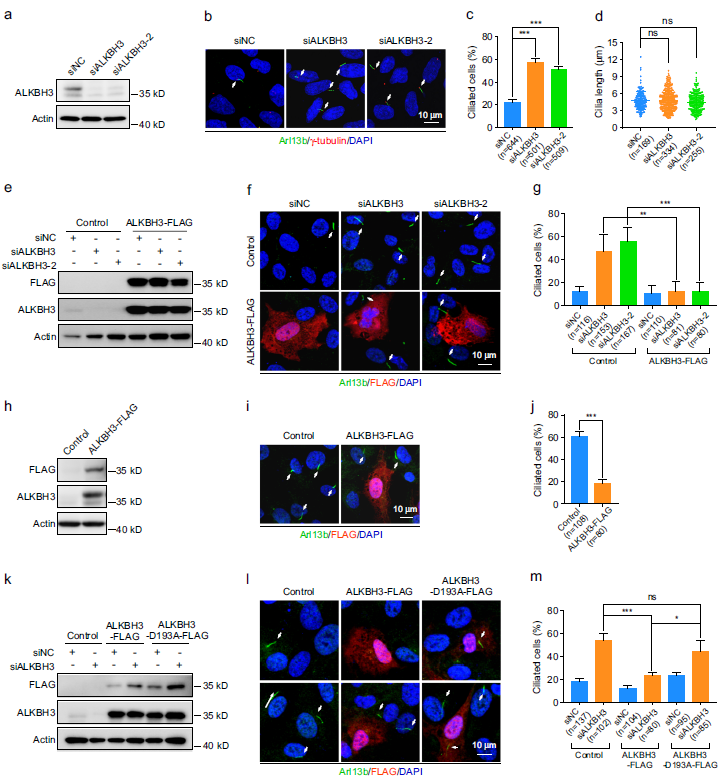
图1:ALKBH3通过其m1A去甲基化活性抑制纤毛发生。
(2) 敲低ALKBH3导致Aurora A mRNA的m1A水平升高
为了探索ALKBH3在纤毛发生中的分子机制,我们试图确定被ALKBH3靶向的可能的mRNAs。首先,我们系统地分析了已发表的HEK-293T细胞中m1A-ID-seq和m1A-quant-seq的数据集,获得了38个潜在的m1A修饰的转录本。这些潜在的m1A修饰的转录本进一步与来自CiliaCarta的纤毛虫相关基因和在缺乏ALKBH3的RPE-1细胞中差异表达的RNA-seq基因重叠。结果,在这四个数据集中,只有Aurora A,一个主要的负调节因子,是重叠的(图2a)。为了研究Aurora A mRNA上是否存在m1A位点,我们进行了基因特异性甲基化RNA免疫沉淀qPCR试验。数据显示,与对照细胞相比,在缺乏ALKBH3的细胞中,Aurora A mRNA的m1A丰度显著增加(图2b),表明Aurora A mRNA可能包含m1A修饰。
为了确定Aurora A mRNA上的精确m1A位点,我们仔细分析了来自野生型和ALKBH3敲除HEK-293T细胞中的m1A-ID-seq数据集,发现在Aurora A mRNA的翻译起始区附近有潜在的m1A峰(图2c)。考虑到RT-1306是一种进化的HIV逆转录酶,能够诱导RNAs上m1A位点的突变,我们使用RT-1306来识别Aurora A mRNA上的m1A位点。我们从对照细胞和ALKBH3缺失细胞的cDNAs中获得的测序数据显示,在Aurora A mRNA翻译起始位点附近的编码序列区存在3个m1A位点(图2d)。此外,我们还将RT-1306扩增的上述cDNAs进行TA克隆,并选取单个菌落进行测序分析。结果显示,在缺乏ALKBH3的细胞中,这些m1A位点的突变率显著增加(图2e)。综上所述,这些观察结果表明,ALKBH3催化Aurora A mRNA上m1A位点的去甲基化。
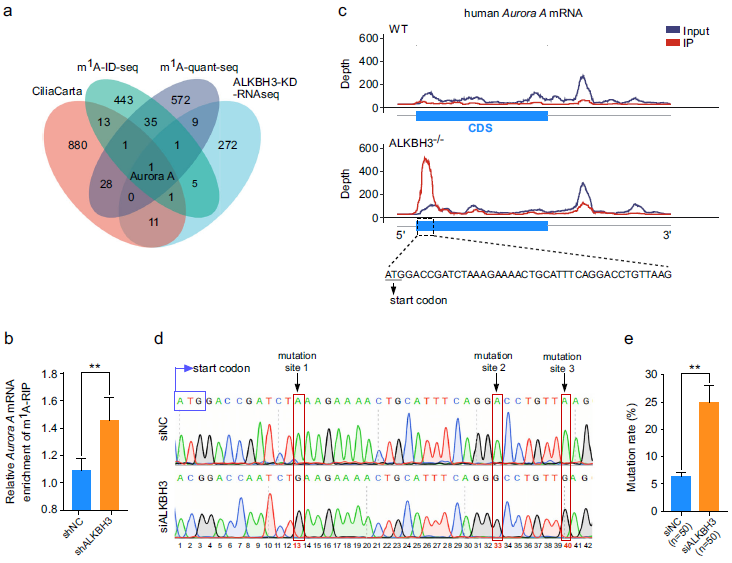
图2:ALKBH3去除Aurora A mRNA上的m1A位点。
(3) ALKBH3促进Aurora A mRNA的稳定和翻译
由于mRNA的m1A修饰被报道影响mRNA的代谢过程,包括mRNA的稳定性和翻译,我们首先研究了ALKBH3是否调节Aurora A的mRNA水平。结果显示,敲低ALKBH3显著降低了RPE-1细胞中Aurora A mRNA的水平(图3a,b)。进一步的实验表明,当细胞中使用放线菌素D暂停转录时,在缺乏ALKBH3的细胞中,Aurora A mRNA的衰减率也显著增强(图3c)。接下来,我们确定了Aurora A mRNA的RNA翻译效率是否受ALKBH3的影响。Western分析表明,ALKBH3的缺失显著降低了Aurora A蛋白水平,而异位表达的ALKBH3有效逆转了这一变化,但突变体ALKBH3-D193A却没有改变(图3d-g)。此外,RPE-1细胞中的多聚体分析显示,稳定敲除ALKBH3会导致高分子量(HMW)多聚体部分的Aurora A mRNA丰度降低,而这些多聚体通常具有较高的翻译效率(图3h,i),这意味着ALKBH3的缺失可能会抑制Aurora A mRNA的翻译。总的来说,这些数据表明,ALKBH3对Aurora A mRNA的稳定性和翻译的调节至关重要。
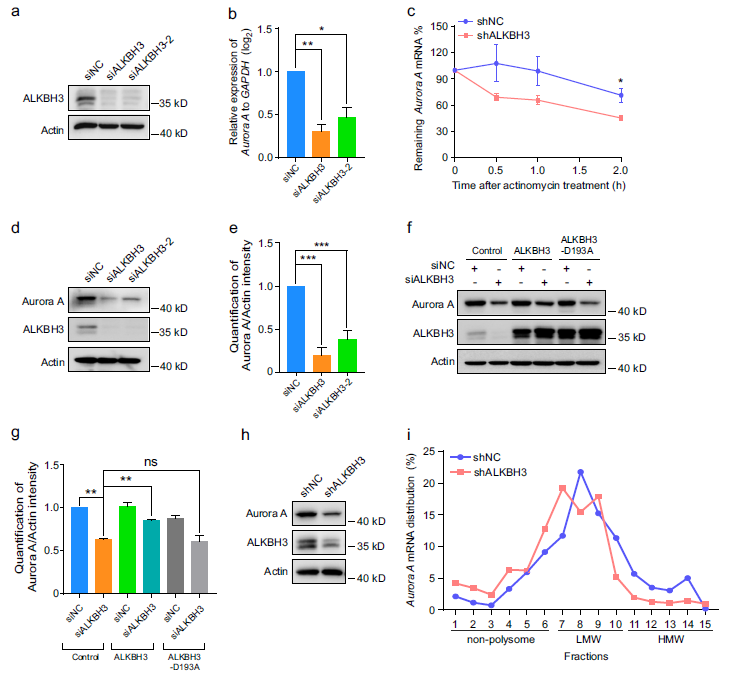
图3:敲低ALKBH3会降低Aurora A mRNA的稳定性和翻译能力。
(4) ALKBH3通过Aurora A抑制纤毛发生
考虑到Aurora A是促进纤毛分解的关键调控因子,我们推断ALKBH3可能影响纤毛分解。为了验证这一点,我们用血清饥饿处理RPE-1细胞24小时以诱导纤毛,然后用血清再刺激24小时以触发纤毛拆卸(图4a)。在此条件下,我们观察到对照组RPE-1细胞在血清刺激24小时后纤毛显著减少(图4b-d)。然而,缺乏ALKBH3的细胞明显抵抗血清诱导的纤毛拆卸。因此,这些数据表明,ALKBH3具有调节纤毛拆卸的功能。我们的数据显示,Aurora A的总水平在血清刺激后2小时略有上升,在18小时和24小时达到峰值(图4e)。我们还发现在血清饥饿/再刺激期间,ALKBH3与Aurora A的表达模式相似(图4e)。值得注意的是,在整个血清刺激过程中,缺乏ALKBH3的RPE-1细胞中的Aurora A蛋白水平明显降低(图4f),这意味着ALKBH3对Aurora A表达的调控可能发生在纤毛周期中。
免疫荧光实验数据显示,在缺乏ALKBH3的细胞中,Aurora A和磷酸化的Aurora A水平都明显降低(补充图未展示)。进一步的结果表明,在缺乏ALKBH3的细胞中,中心体上Aurora A和磷酸化的Aurora A的荧光强度显著降低(补充图未展示)。中心体上Aurora A和磷酸化的Aurora A的细胞比例也显著降低(补充图未展示)。这些观察结果表明,ALKBH3的消耗降低了中心体Aurora A和磷酸化Aurora A的水平。此外,Aurora A的沉默显著促进了RPE-1细胞的纤毛发生(补充图未展示)。拯救实验表明,外源表达GFP-Aurora A蛋白能够显著逆转由ALKBH3敲低引起的增强的纤毛(图4g-i)。这些数据共同表明,ALKBH3通过促进Aurora A的表达来抑制纤毛。
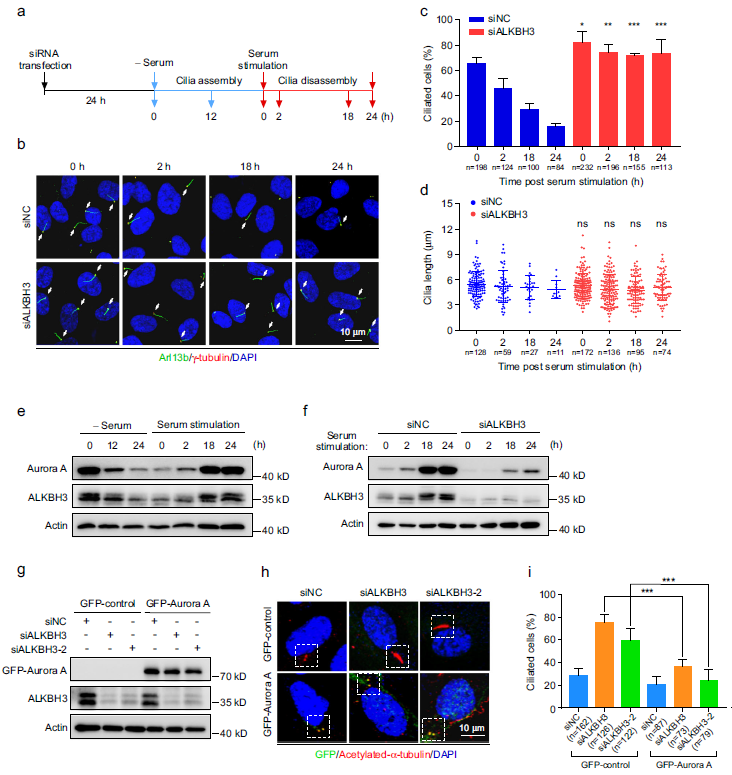
图4:Aurora A的异位表达逆转了ALKBH3缺失细胞的纤毛缺陷。
(5) ALKBH3缺乏促进纤毛诱导细胞周期阻滞
为了探索缺乏ALKBH3的细胞中增强的纤毛是否与细胞周期阻滞相关,我们首先检测了无血清饥饿的RPE-1细胞中缺乏ALKBH3对细胞周期进展的影响。FACS分析显示,ALKBH3缺失增加了G0/G1期细胞的百分比(补充图未展示)。免疫印迹分析显示,在ALKBH3缺失的细胞中,细胞周期标志物cyclin A减少,表明在ALKBH3敲低的细胞中,S期或G2/M期细胞减少(补充图未展示)。进一步的EdU-标记实验显示,当ALKBH3耗尽时,EdU-结合细胞的百分比显著下降(补充图未展示)。这些数据表明,ALKBH3的耗尽导致细胞周期在G0/G1期停止。然后,我们研究了由ALKBH3耗竭引起的G0/G1期细胞周期阻滞是否由其增强的纤毛介导。来自FACS谱、细胞周期标记和EdU合并的数据显示,IFT20耗尽诱导的纤毛脱落显著逆转了ALKBH3耗尽细胞中G0/G1阻滞(补充图未展示),表明ALKBH3耗尽以纤毛依赖的方式诱导G0/G1阻滞。
接下来,我们研究了Aurora A敲除是否也会导致RPE-1细胞中纤毛依赖的细胞周期阻滞。Aurora A的沉默会导致细胞周期在G0/G1期停止,同时IFT20的耗尽也会显著逆转(补充图未展示)。总的来说,这些结果表明,无论是ALKBH3还是Aurora A的缺失都会导致RPE-1的纤毛增强,这可能随后导致细胞周期在G0/G1期停止。
(6) 斑马鱼ALKBH3的分子特征
为了探索ALKBH3在脊椎动物发育过程中纤毛发生中的作用,我们首先克隆了斑马鱼ALKBH3(图5a)。生物信息学分析显示,ALKBH3编码一个推导出的282 aa蛋白,具有保守的2OG-Fe(II)加氧酶超家族域,这意味着斑马鱼的ALKBH3也可能具有去甲基化活性(图5b)。比对分析表明斑马鱼ALKBH3与人类ALKBH3具有高度同源性(图5c)。然后,我们检测了斑马鱼ALKBH3在胚胎发育过程中的时间表达,发现ALKBH3 mRNA由母体沉积,并在整个早期斑马鱼胚胎阶段普遍表达(图5d)。全安装原位杂交显示,ALKBH3 mRNA由母体提供,在48 hpf(受精后小时)到72 hpf时在大脑中富集,在96 hpf时下降到低水平(图5e)。
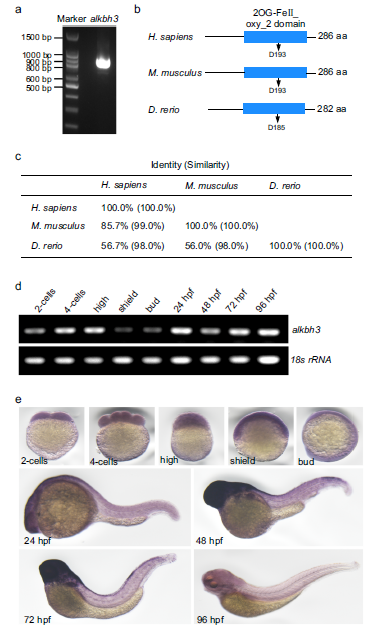
图5:斑马鱼ALKBH3基因的分子特征。
(7) ALKBH3突变体在斑马鱼胚胎中显示纤毛表型
为了研究ALKBH3是否影响斑马鱼的绒毛发生和胚胎发育,我们设计了反义吗啉代寡核苷酸(ALKBH3 MO)来阻断ALKBH3 mRNA的翻译。ALKBH3突变体在斑马鱼胚胎中显示出纤毛表型,包括身体弯曲、心包水肿、耳囊泡内异常耳石和前肾管扩张(图6a-d)。重要的是,通过与ALKBH3 mRNA共注射,ALKBH3突变体中的这些缺陷被显著逆转,但根据氨基酸序列的进化保守分析,预测到催化活性不强的ALKBH3突变体的ALKBH3-D185A mRNA没有被逆转(图5b)。进一步的观察表明,在28 hpf时,与对照变体相比,ALKBH3突变体的前肾管纤毛明显有缺陷,异位表达的ALKBH3 mRNA而不是异位表达的ALKBH3-D185A mRNA也显著挽救了纤毛(图6e,f)。总之,这些数据表明,ALKBH3可能通过其在斑马鱼体内的去甲基化活性在纤毛相关的发育过程中发挥关键作用。
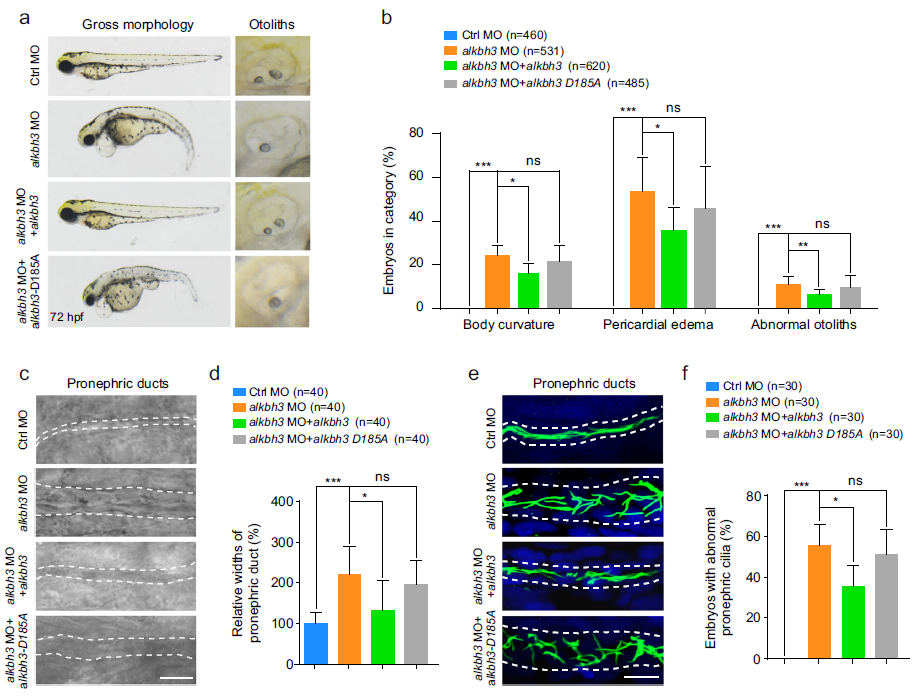
图6:斑马鱼ALKBH3突变体表现出纤毛表型。
(8) Aurora A的异位表达逆转了ALKBH3突变体的纤毛缺陷
为了确定Aurora A是否参与斑马鱼胚胎发育过程中ALKBH3对纤毛相关事件的调控,我们检测了ALKBH3突变体中Aurora A mRNA的表达水平,发现Aurora A mRNA在缺乏ALKBH3的胚胎中显著降低(图7a),这与我们从哺乳动物细胞中获得的数据一致(图3b)。此外,共注射Aurora A mRNA能够显著逆转ALKBH3突变体的纤毛表型(图7b-e)。其他数据显示,异位表达的Aurora A mRNA显著挽救了ALKBH3敲除胚胎前肾导管的异常纤毛(图7f,g)。总之,这些结果表明,ALKBH3可能通过调节脊椎动物胚胎发生中的Aurora A mRNA来影响纤毛相关的发育事件。
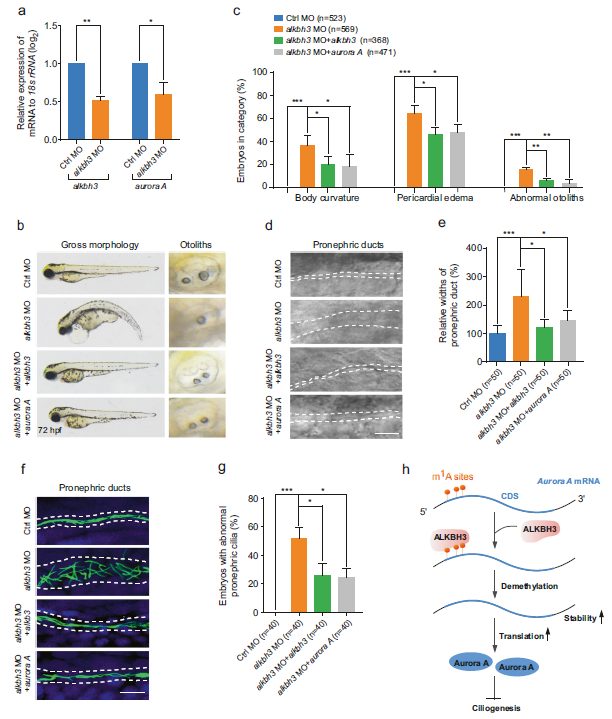
图7:Aurora A的异位表达挽救了ALKBH3形态的纤毛缺陷。
结论:
我们的研究揭示了m1A去甲基化酶ALKBH3在脊椎动物纤毛发生和胚胎发育中先前未被描述的作用。ALKBH3在翻译起始位点附近的编码序列区去除Aurora A mRNA的m1A修饰,抑制Aurora A mRNA的衰变并促进其mRNA的翻译,从而维持Aurora A蛋白的丰度以抑制纤毛发生(图7h)。
参考文献:
Kuang, W., Jin, H., Yang, F., Chen, X., Liu, J., Li, T., Chang, Y., Liu, M., Xu, Z., Huo, C., Yan, X., Yang, Y., Liu, W., Shu, Q., Xie, S., & Zhou, T. (2022). ALKBH3-dependent m1A demethylation of Aurora A mRNA inhibits ciliogenesis. Cell discovery, 8(1), 25. https://doi.org/10.1038/s41421-022-00385-3.
