






LnRNA提供了重要的染色质支架——为蛋白质相互作用和骨髓瘤生长
多发性骨髓瘤是一种复杂的浆细胞恶性肿瘤,约占血液系统肿瘤的10%,目前仍无法治愈。越来越多的证据表明非编码RNA(noncoding RNA,ncRNA)在多发性骨髓瘤中发挥关键作用。在本研究中,作者使用了大规模的CRISPR干扰活性筛选来询问多发性骨髓瘤中细胞生长对lncRNA基因的依赖性,并确定了miR-17-92集群宿主基因(MIR17HG)的突出作用,发现一种MIR17HG衍生的lnc-17-92,以一种独立于microRNA和DROSHA的方式为c-MYC与WDR82之间的相互作用提供了染色质支架,从而促进ACACA的表达。本研究建立了一种新的致癌基因MIR17HG,为转化临床试验提供了有效的抑制剂。
技术路线:
主要研究结果:
1、CRISPRi活性筛选确定MIR17HG为MM中主要的细胞生长依赖
作者分析了360个新诊断的多发性骨髓瘤(MM)患者的RNA-seq数据,并在原代MM细胞和70个MM细胞系中鉴定了913个lncRNA转录本( 图1A,I )。为系统地研究这些lncRNA在MM细胞生长中的作用,作者转导了3个MM细胞系( H929、KMS-11和KMS-12-BM ),该细胞系表达了dCAS9 - KRAB融合蛋白,该文库由7个sgRNA(small guide RNA)组成,分别针对所识别的lncRNA的913个转录起始位点( TSS )和576个阴性对照sgRNA ( 图1A、II )。3周后,作者使用深度测序和基于模型的全基因组CRISPR - Cas9基因敲除( MAGeCK )稳健排序聚合算法( RRA )对MM细胞群体中相对贫乏或富集的sgRNA进行了测试。
最富集或最贫乏的sgRNA在次级筛选中进一步测试,使用224个靶向lncRNA的TSS混合文库,已知蛋白编码癌基因( MYC、IRF4 )或肿瘤抑制因子( TP53 )的TSS作为阳性对照,2245个非靶向sgRNA作为阴性对照( 图1A,III )。在次级筛选中,4个MM细胞系( H929、KMS11、KMS12BM和AMO1)被用来检测和排序显著耗尽或富集的sgRNA。如预期的那样,靶向IRF4和MYC的sgRNA在3种( MYC )或全部( IRF4 )细胞系中显著缺失,而靶向TP53的sgRNA在两种TP53野生型细胞系( AMO1和H929 )中显著富集。
作者对sgRNA敲除进行排序分析,发现MIR17HG是最主要的依赖,在所有测试的细胞系中,RRA评分等于或优于靶向MYC或IRF4的评分(图1B)。为进一步验证这一数据,作者接下来在四环素诱导系统启动子的调控下,转导4个靶向MIR17HG的sgRNAs转导表达dCAS9 - KRAB融合蛋白的MM细胞系,观察到感染非靶向sgRNAs的细胞相比,持续暴露于强力霉素后,细胞生长减少(图1C)。此外,作者使用2种不同的锁核酸( LNA ) gapmeR ASOs(反义寡核苷酸)靶向MIR17HG新生RNA ( pre-RNA ),用于RNase H介导的降解,转染11种MM细胞系,包括对常规抗MM药物(AMO1-ABZB对硼替佐米耐药;AMO1-ACFZ对卡非佐米耐药;MM.1 R对地塞米松耐药)耐药的细胞系,并证实对MM细胞活力的显著影响独立于遗传和分子背景(图1D)。
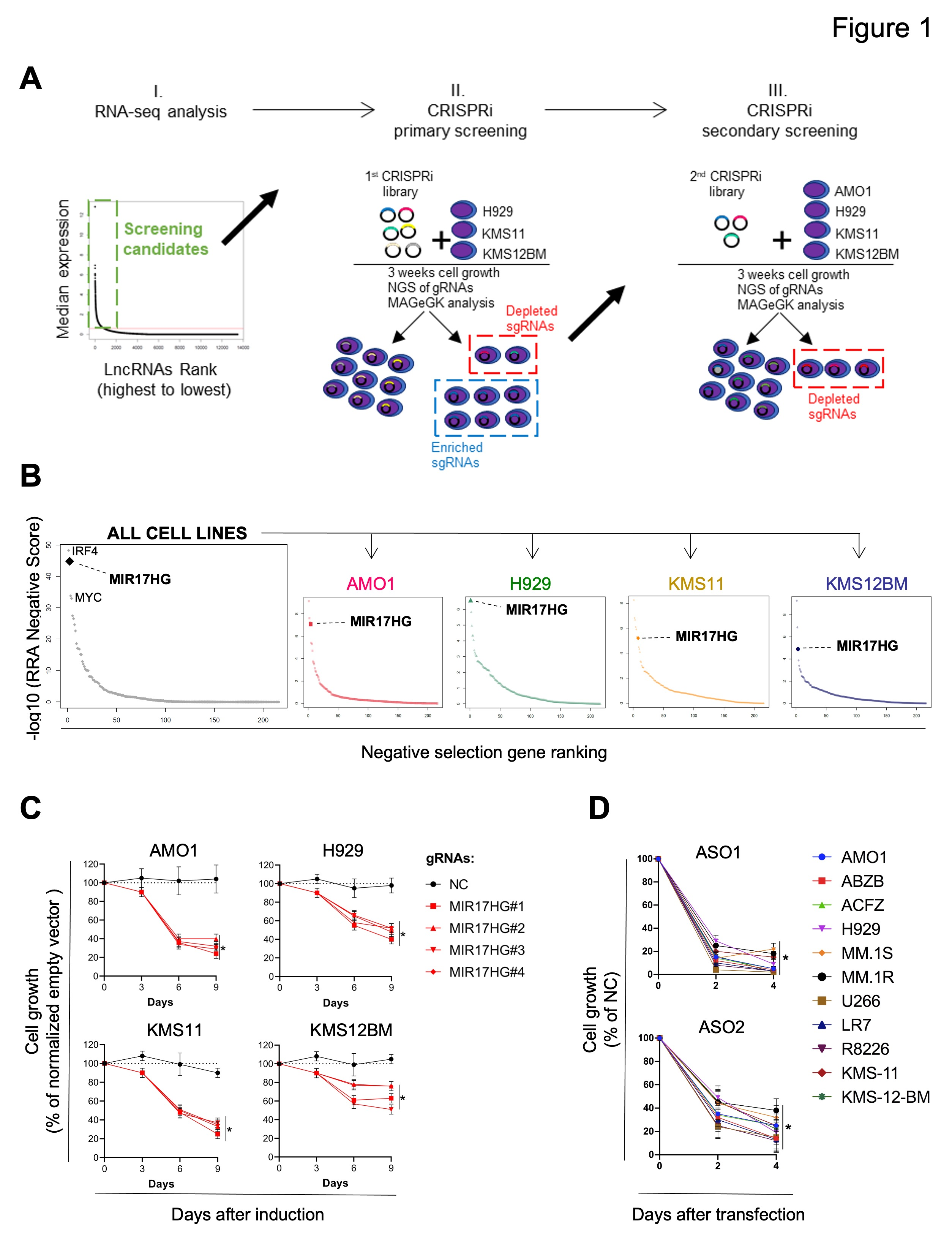
图1 CRISPRi活性筛选确定MIR17HG为MM中主要的细胞生长依赖
2、MIR17HG衍生的lnc-17-92以不依赖于miRNA和DROSHA的方式介导细胞生长依赖性
MIR17HG是miRNA簇miR-17-92和lncRNA的一个位点,命名为lnc-17-92。lnc-17-92有两种异构体,一种是~ 5000 nt长的(lnc-17-92TV1),另一种是~ 900 nt的( lnc-17-92TV2),它们都尚未被功能研究( 图2A )。在MM细胞系中,作者通过单分子RNA-FISH证明了lnc-17-92的核富集( 图2B )。作者发现在3个大队列的初诊MM患者中,lnc-17-92的高表达与更短的无事件生存( EFS )和总生存( OS )相关( 图2C )。
为检测lnc-17-92的独立活性,作者首先通过异位表达原始前体pri-mir-17-92建立过表达miR-17-92的两种MM细胞系。在这些细胞系中特异性敲除lnc-17-92,ASOs靶向MIR17HG前体RNA的5’ 端,该区域不被pri-mir-17-92覆盖,并观察到细胞生长的显著抑制,而异位的pri-mir-17-92不能挽救细胞生长( 图2D )。接下来,作者建立了两个DROSHA基因敲除( DR-KO )的MM细胞系( AMO1DR-KO和H929DR-KO),使用ASO1处理敲低lnc-17-92后,在DR-WT和DR-KO细胞系统中仍然观察到强烈的抗增殖活性( 图2E )。重要的是,在NOD SCID小鼠中,暴露于gymnotic ASO1会破坏AMO1DR-KO细胞建立肿瘤的能力,导致动物生存期延长(图2E - G)。这些结果表明,lnc-17-92TV1是MIR17HG癌症依赖性的主要中介因子,独立于miR-17-92的作用和生物发生途径。

图2 MIR17HG衍生的lnc-17-92以不依赖于miRNA和DROSHA的方式介导细胞生长依赖性
3、Lnc-17-92与ACACA形成转录轴,促进MM细胞生长
Lnc-17-92的核富集提示其可能在基因表达调控中发挥作用。因此,作者在DR-WT ( AMO1和H929 )和DR-KO ( AMO1DR-KO ) MM细胞系中使用早期暴露于gymnotic ASO1来消除lnc-17-92,以避免在DROSHA WT细胞中调节miR-17-92和miR-17-92的经典靶标,并在所有测试的细胞系中鉴定出7个lnc-17-92消除后迅速下调的基因( 图3A )。作者在体外用ASO1处理3例MM患者的CD138 +细胞验证了这些发现( 图3B )。此外,作者在2个大型RNA-seq MM患者数据集( IFM / DFCI和MMRF / CoMMpass)中观察到lnc-17-92与其靶基因之间存在显著的正相关性( Spearman r > 0.3 ; p < 0.001) ( 图3C )。
利用荧光素酶报告基因实验,在293TDR - KO细胞中,在存在或不存在lnc-17-92缺失的情况下,作者证明了lnc-17-92对这些基因的调控,除了ANO6,发生在启动子水平( 图3D )。与此一致,作者通过染色质分离RNA沉淀( ChIRP )实验和qRT - PCR分析证实了lnc-17-92在顶端靶标ACACA的启动子区域存在相互作用( 图3E ),并且通过对lnc-17-92和ACACA前体mRNA的单分子双RNA FISH分析显示lnc-17-92频繁地定位到ACACA位点( 图3F )。与随机斑点相比,lnc-17-92在ACACA基因位点的近端定位明显更频繁( 图3F )。
在已确定的lnc-17-92靶点中,ACACA对MM细胞的增殖和存活影响最大( 图3G )。ACACA编码脂肪从头合成途径的限速酶ACC1,在不同的癌症环境中支持肿瘤发生。这些数据表明lnc-17-92是一种染色质相互作用的lncRNA,具有转录调节功能。
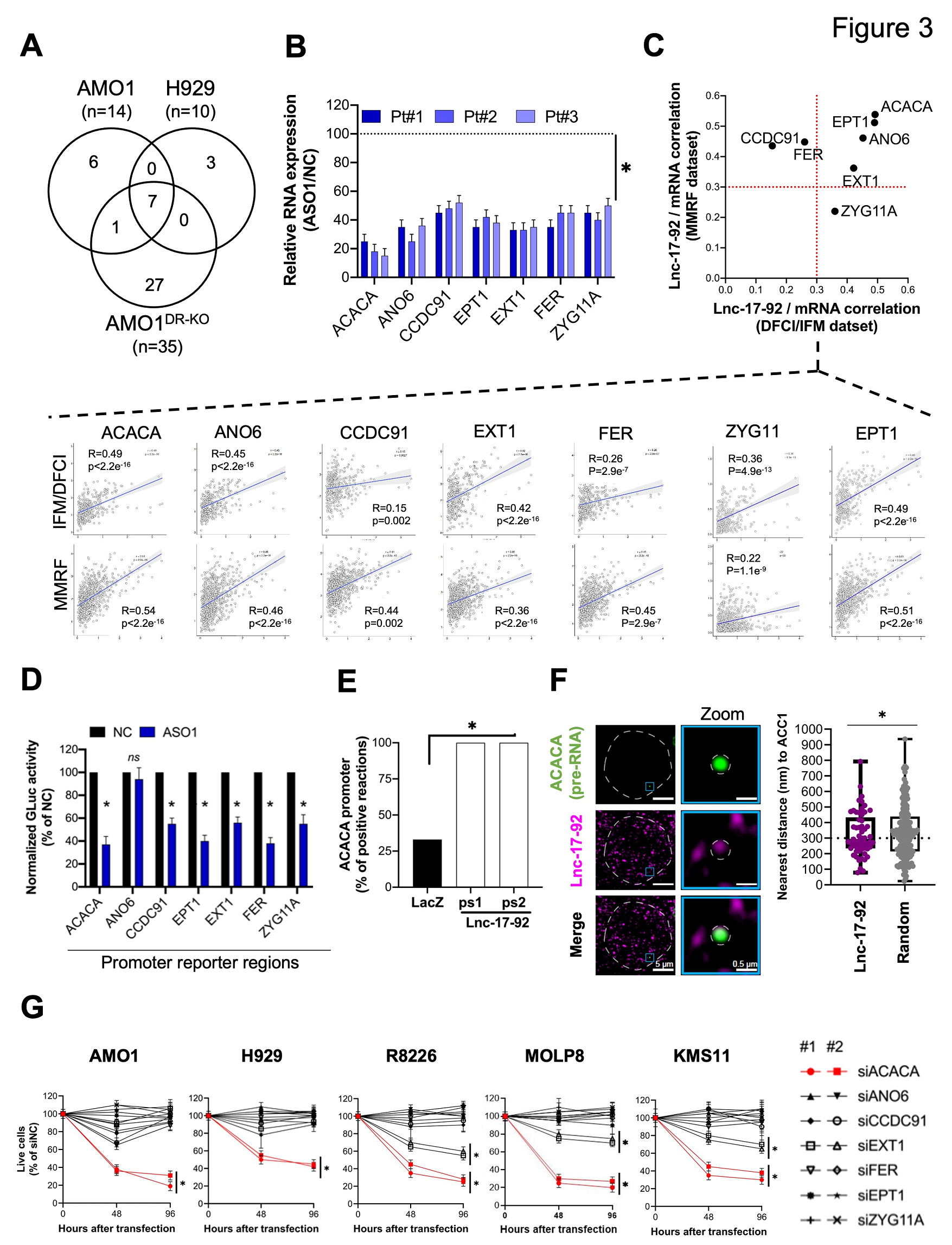
图3 Lnc-17-92与ACACA形成转录轴,促进MM细胞生长
4、Lnc-17-92直接与c-MYC相互作用,促进其在ACACA启动子上的占用。
作者进行RNA Protein pull-down ( RPPD )实验,发现MYC与lnc-17-92TV1形成复合物( 图4A )。用MYC抗体进行RNA免疫沉淀实验( RIP )证实lnc-17-92TV1的富集( 图4B )。此外,RNA yeast-3-杂交( Y3H )实验证实了在体内细胞模型中lnc-17-92TV1-MYC相互作用,如酵母克隆生长所示( 图4C )。
接下来,作者评估MYC和lnc-17-92是否协同促进ACACA在MM细胞中的表达。在MM细胞中敲除lnc-17-92的确可以在不影响MYC表达的情况下消除MYC在ACACA启动子上的占位( 图4D ),并且仅在高MYC水平存在的条件MYC Tet - Off细胞系P493 - 6中降低ACACA的表达( 图4E )。这些数据表明lnc-17-92TV1与转录因子MYC形成RNA -protein复合体,促进其在ACACA启动子上的染色质占用和转录活性。

图4 lnc-17-92直接与c-MYC相互作用,促进其在ACACA启动子上的占用
5、Lnc-17-92介导MYC-WDR82转录复合体的组装,导致ACACA的转录和表观遗传激活
为确定lnc-17-92是否影响这些蛋白质-蛋白质相互作用,作者在三个MM细胞系( AMO1、H929和U266MYC +)中整合了邻近依赖性生物素识别( BioID )分析、免疫共沉淀实验和质谱分析( Co-IP/MS )的结果。这一分析突出了WDR82是一个非常高可信度的lnc-17-92依赖的MYC互作因子( 图5A )。RPPD和RNA Y3H实验进一步证实lnc - 17 - 92TV1和WDR82之间存在直接的RNA -protein相互作用( 图5B-C )。
WDR82是SET1甲基转移酶复合物的调节成分,它在活性位点的转录起始位点催化组蛋白H3 Lys-4 ( H3K4 )甲基化( mono-,di-,tri-),这是MYC与染色质结合和反式激活的先决条件。在MM细胞中,敲低WDR82降低了ACACA启动子上H3K4me3和MYC的占位(图5D-E),降低了ACACA mRNA表达(图5F)。这证实在MM细胞中沉默WDR82对H3K4甲基化的整体影响。此外,研究利用表达异位WDR82 - GFP融合蛋白的MM细胞证明了lnc-17-92表达对于WDR82在ACACA启动子上的占位是必需的(图5G)。此外,lnc-17-92缺失导致ACACA启动子上H3K4me3水平降低(图5H),但不影响H3K4甲基化状态( 图5I )。
这些发现表明lnc-17-92TV1是一个染色质支架,介导MYC-WDR82多蛋白转录复合体的组装,以控制ACACA和可能的其他基因的表达。

图5 Lnc-17-92介导MYC-WDR82转录复合体的组装,导致ACACA的转录和表观遗传激活
6、MIR17HG的治疗抑制剂在人MM动物模型体内外均具有抗肿瘤活性
为开发临床应用的抑制剂,作者筛选了八十多个fully phosphorothioated ( PS )、2 ' – O-methoxyethyl(2 '-MOE)修饰、lipid-conjugated的ASOs,它们既可以触发RNase H介导的MIR17HG pre-RNA ( gapmeRs ) 降解,也可以通过RNase H-independent机制( blockmeRs )发挥作用。该方法鉴定了一种18-mer tocopherol (T)-conjugated gapmeR G2-15b-T ( ' G ' )和一种18 – mer tocopherol (T)-conjugated steric blockers SB9-19-T ( ' B ' ),它们在大量MM细胞系和CD138 +原代MM细胞中都具有强烈的抗增殖作用,同时保留了来自3个健康捐献者的非恶性细胞系(THLE-2、HK-2、HS-5和293T)和PBMC(外周血单核细胞)。
为评估这两个化合物的体内抗肿瘤活性,作者首先使用免疫缺陷的NOD SCID小鼠建立了基于AMO1的浆细胞瘤异种移植模型。在这里,作者观察到G2-15b-T(肿瘤生长抑制,TGI=76%)或B9-19-T(TGI=69%)治疗周期后肿瘤生长显著减少(图6A)。对该治疗后小鼠的肿瘤进行分析,证实了lnc-17-92的表达降低(图6B),以及lnc-17-92的靶标( ACACA、EPT1、EXT1、CCDC91、ANO6、FER和ZYG11A)的调节(图6C)。
接下来在弥漫性骨髓瘤侵袭模型中证实了G2-15b-T和B9-19-T具有显著的抗MM活性,其中MOLP8-luc + MM细胞的肿瘤生长通过生物发光成像( BLI )评估。在该模型中,用G2-15b-T ( TGI = 84 % )或B9-19-T ( TGI = 52 % )治疗一个周期后,肿瘤生长明显被拮抗。用G2-15b-T治疗后,8只小鼠中有2只( 25 % )肿瘤被清除(图6D)。重要的是,两种抑制剂都显著延长了动物的存活时间(图6E)。
最后,作者通过尾静脉注射从晚期患者( PDX-NSG )获得的CD138 + MM细胞,建立了临床相关的PDX-NSG小鼠模型。在这个模型中,使用人类kappa light chain作为替代物在血清样本中监测肿瘤生长。值得注意的是,作者观察到G2-15b-T治疗周期后肿瘤生长的消退,其效果与硼替佐米(阳性对照)相当(图6F)。
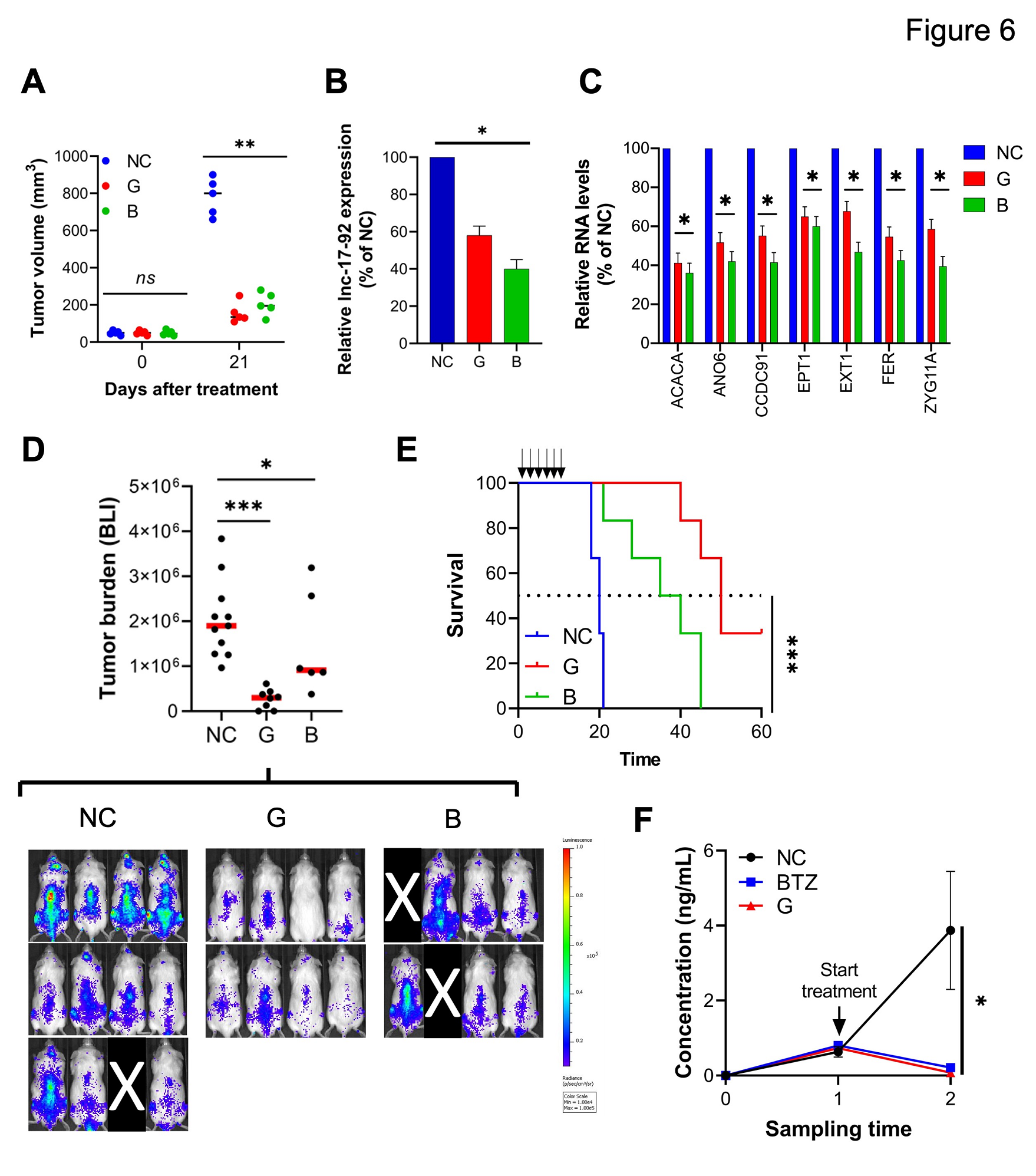
图6 MIR17HG的治疗抑制剂在人MM动物模型体内外均具有抗肿瘤活性
结论
总的来说,这项研究建立了具有独特的lncRNA功能的MIR17HG,它能促进蛋白质-蛋白质和蛋白质-DNA相互作用,对促进肿瘤治疗有积极意义。
参考文献
Morelli E, Fulciniti M, Samur MK, Ribeiro C, Wert-Lamas L, Henninger JE, et al. A MIR17HG-derived Long Noncoding RNA Provides an Essential Chromatin Scaffold for Protein Interaction and Myeloma Growth. Blood. 2022 Sep 20:blood.2022016892. doi: 10.1182/blood.2022016892.