






脂肪多糖衍生亚油酸促进米色脂肪祖细胞增殖
新生米色脂肪细胞的生物发生涉及白色脂肪组织(WAT)中祖细胞的增殖;然而,是什么调控了这一过程仍不清楚。在这里,本演技报告了在小鼠模型和人类组织中,WAT脂解衍生的亚油酸在冷适应、b3-AR激活和烧伤后触发米色祖细胞增殖。细胞表面标记物PDGFRa或Sca1和CD81标记的脂肪祖细胞亚群含有丰富的嵴线粒体,并通过脂肪酸转运蛋白CD36主动输入亚油酸。口服补充亚油酸,即使在热中性条件下以CD36依赖的方式刺激米色祖细胞增殖。总之,本研究结果为多样化的病理生理提供机制性的见解。本研究于2022年12月发表于期刊《Developmental cell》上,IF:1.417。
技术路线:
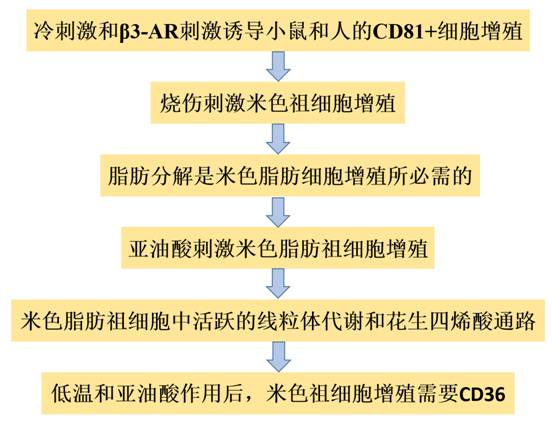
主要研究结果:
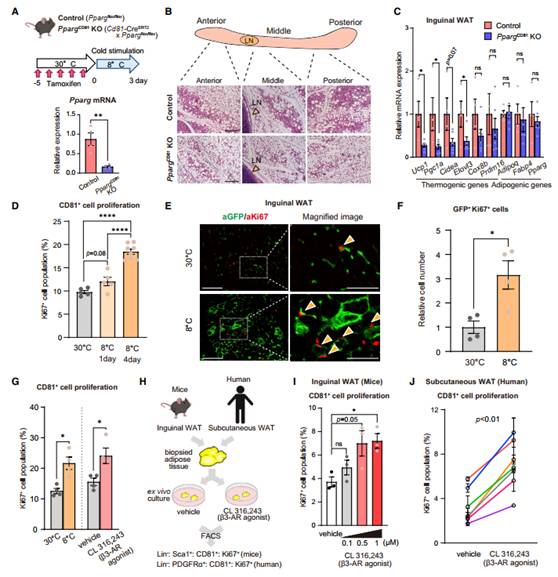
作者建立在他莫昔芬处理后的CD81 +细胞中选择性缺乏脂肪生成主要调节因子PPARg的小鼠。为此,作者在PpargCD81 KO小鼠中建立了他莫昔芬治疗后在CD81 +细胞中选择性缺乏脂肪生成的主要调节因子PPARg的小鼠(图1A)。小鼠在30℃下接受他莫昔芬治疗,随后适应8℃。冷刺激下,对照组小鼠(Ppargflox/flox)在腹股沟WAT(白色脂肪组织)的前、中、后区有大量表达UCP1(解偶联蛋白1)的多室脂肪细胞簇,这是米色脂肪细胞的一个形态学特征(图1B)。相比之下,PpargCD81 KO小鼠腹股沟WAT在冷刺激下白色脂肪组织大大受损(图1B)。与形态学变化一致,PpargCD81 KO小鼠腹股沟WAT中棕色/米色脂肪选择基因UCP1、Pgc1a、Cidea和Elovl3的表达水平明显低于对照组小鼠(图1C)。这些结果说明大量冷刺激诱导下的白色脂肪组织起源于小鼠腹股沟WATCD81+祖细胞(APC)。作者发现腹股沟WAT中CD81 +APC(Lin- : Sca1 + : CD81 + : Ki67 +)在冷暴露3天内Ki67 +细胞群数量显著增加(图1D)。用CD81-lineage reporter小鼠进行组织学分析也表明腹股沟WAT的基质细胞共表达GFP和Ki67,Ki67+ GFP+细胞的数量在冷刺激下显著增长(图1E-F)。除了冷刺激,用b3-AR治疗3天,相较于载体处理小鼠WAT,治疗后小鼠腹股沟WAT中Ki67+和CD81+细胞增多(图1G)。FACS(荧光激活细胞分选术)定量地检测来自活检组织Lin svf中Ki67+和CD81+细胞的数量,发现CL316,243在体外培养的小鼠腹股沟WAT中以剂量依赖的方式显著增加Ki67+和CD81+细胞的数量(图1I)。在人WAT中b3-AR治疗也显著增加Ki67+和CD81+细胞的数量,尽管处于非刺激下的Ki67+和CD81+细胞数量与个体之间的有所不同(图1J)。
根据在小鼠中建立的方案,在烧伤和假手术处理后,作者将获得腹股沟WAT 1天、3天和7天的C57BL/6J小鼠(图2A)。烧伤后3天和7天血浆游离脂肪酸(FAA)水平明显高于对照组小鼠(图2B)。烧伤后7天,受伤小鼠腹股沟WAT各区域均有大量多房型米色脂肪细胞,其中大部分表达UCP1蛋白(图2C)。与对照组小鼠相比,在烧伤后7天,形态变化伴随着棕/米色选择基因的表达增加,如Ucp1、Elovl3、Dio2和Cox8b(图2D)。重要的是,作者发现在烧伤3天和7天后,小鼠腹股沟WAT中Ki67+CD81+细胞显著增多(图2E-F)。这些结果表明米色脂肪发生的病理刺激包括WAT中CD81+祖细胞的增殖。
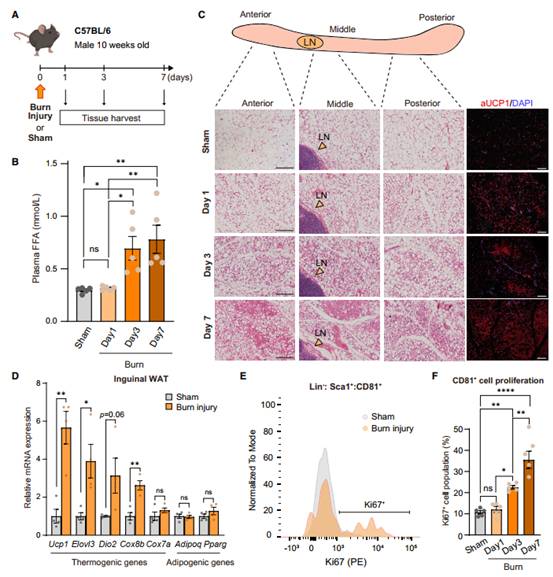
图2 烧伤促进CD81+ APC细胞增殖
3、脂肪分解是米色脂肪细胞增殖所必需的
为验证猜想WAT脂肪多聚物衍生因子介导米色祖细胞的增殖,作者使用缺乏脂肪肝三酯脂肪酶(ATGL)小鼠,即Adipo-ATGL KO小鼠。在这个实验中,Adipo-ATGL KO小鼠和它的同窝出生的小鼠在30℃下生存14天,随后8℃下生存3天(图3A-B)。
冷驯化3天后,对照组小鼠腹沟WAT库的组织学分析发现大量米黄色脂肪细胞,多房脂质和UCP1表达,特别是在中间区域。相反,在AdipoATGL KO小鼠中,冷诱导的米色脂肪细胞的生物生成几乎完全消失(图3C)。此外,adipocyte-specificATGL缺失显著降低了在腹股沟WAT中棕色/米色脂肪选择基因的表达,包括Ucp1、Pgc1a、Cidea、Dio2、Cox8b和Prdm16(图3D)。
接着FACS检测30C和冷驯化后小鼠腹股沟WAT中CD81+细胞的增殖。30℃时,Ki67+ CD81+细胞数量在基因型间无差异。冷驯化显著增加对照组小鼠Ki67+ CD81+细胞数量;然而,这种冷诱导的米色祖细胞增殖在Adipo-ATGL KO小鼠中完全消失(图3E)。当CL316243处理体外培养的腹股沟WAT时,观察到对照小鼠来源的WAT库中的CD81+ Ki67+细胞的数量显著增加。相比之下,CL316243在AdipoATGL KO小鼠中的刺激作用减弱(图3F)。与遗传学研究一致的是,Atglistatin对ATGL的药理抑制有效的阻断CL316243对野生型小鼠腹股沟WAT中CD81+细胞增殖的刺激(图3G)。
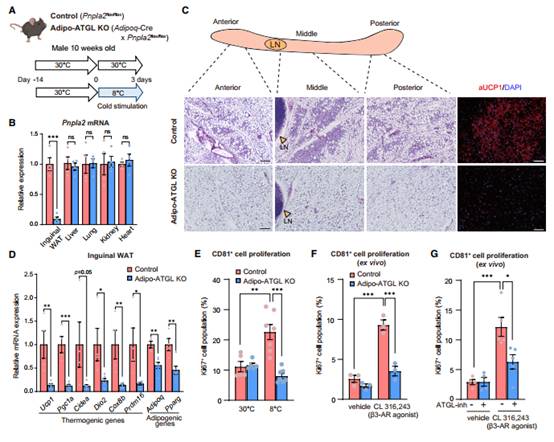
图3 CD81+ APC细胞增殖需要脂解作用
4、亚油酸刺激米色脂肪祖细胞增殖
作者下面鉴定脂质分解因素介导的米色脂肪细胞增殖(图4A)。煮沸培养基中的脂质提取物能够刺激CD81+细胞增殖(图4B)。随后,作者基于前人的数据库和质谱循环分析证明了冷刺激和b3-AR治疗都能持续的增加,如图4C所示的6种脂肪酸。补充亚油酸显著刺激CD81+细胞的增殖,而其他细胞没有明显变化(图4D)。亚油酸处理通过增加S期和G2-M期细胞数量促进细胞增殖,CD81+细胞优先增殖(图4E)。同时,作者也发现是亚油酸,而不是a亚麻酸或反亚麻酸,亚油酸的几何异构体促进CD81+细胞增殖(图4F)。为检测口服亚油酸是否促进体内米色细胞增殖,野生型雄性小鼠在30℃条件下,每天摄入浓度为1%的亚油酸(图4G)。FACS检测发现口服亚油酸可使小鼠体内Ki67+ CD81+细胞显著增加3倍(图4H)。尽管8℃的冷刺激会促使处理过的小鼠米色脂肪的生物生成,但是补充亚油酸进一步促进小鼠米色脂肪的生物生成(图4I)。作者发现补充亚油酸增强腹股沟WAT中CD81+细胞米色脂肪的生成(图4J)。作者发现补充亚油酸可以有效刺激对照组中小鼠腹股沟WAT中米色脂肪的生成,而在PpargCD81 KO小鼠中这种作用明显减弱(图4K)。与分子分析一致的是,腹股沟WAT的组织学分析显示PpargCD81KO小鼠的多房米色脂肪细胞比对照组少(图4L)。这些结果表明,当补充亚油酸和冷刺激结合时,增强CD81+细胞来源的新生米色脂肪的生物生成。

图4 亚油酸刺激CD81+APC增殖
5、米色脂肪祖细胞中活跃的线粒体代谢和花生四烯酸通路
为探索亚油酸刺激米色祖细胞增的机制,作者对腹股沟WAT中原生CD81+和CD81-细胞进行RNA测序。与之前的分析一致,转录组学分析显示,与CD81-细胞相比,CD81+细胞表达出高水平的平滑肌基因富集,包括Acta2、Sm22和Myh11(图5A)。值得注意的是,CD81+细胞表达的线粒体编码基因,如Atp6、Atp8、Cox1、Cox2和Cytb,明显高于来自腹股沟WAT的CD81-细胞(图5A)。电子显微镜(EM)中发现CD81+细胞中含有大量的线粒体,其中许多线粒体呈球形或椭圆形,并平行于致密嵴,表现出棕色前脂肪细胞的形态学特征(图5B)。此外,CD81+细胞的耗氧率明显高于基础状态和羰基氰化物4-(三氟甲氧基)苯腙(FCCP)处理后的CD81细胞(图5C)。补充亚油酸通过增强FA氧化进一步激活CD81 +细胞的生物能量学,因为肉碱棕榈酰转移酶(CPT)抑制剂(乙莫克害)抑制FA氧化减弱亚油酸对细胞呼吸的刺激作用(图5D)。
从转录组数据中发现CD81+细胞与CD81细胞相比,花生四烯酸合成途径高度富集(图5E)。接下来,脂质组学检测暴露8℃ 3天的小鼠原代CD81+和CD81-细胞中前列腺素的水平。分析发现PGD2在CD81+细胞中的水平显著高于CD81-细胞(图5F)。随后,液相色谱-串联质谱法(LC-MS / MS)定量CD81+细胞中PGD2的含量以及释放到培养基中的PGD2水平,发现抑制Cox2显著降低PGD2的细胞水平和释放水平(图5G),表明Cox2是CD81+细胞合成PGD2所必需的。Cox1、Alox5和Alox12的抑制剂不干扰亚油酸的作用(图5H)。其次,鉴于CD81+细胞表达高水平的前列腺素受体,如DP1,作者确定PGD2直接作用于米色祖细胞的程度。另外还发现PGD2可以显著促进CD81+细胞的增殖,而DP1的特异性抑制剂MK-0524可以阻断PGD2的促增殖作用(图5I)。冷暴露显著增加腹股沟WAT中Ki67 + CD81 +细胞的数量;然而,Cox2抑制剂处理显著减弱冷暴露对CD81 +祖细胞增殖的刺激作用(图5J)。这些结果表明脂解衍生的亚油酸被线粒体中的CD81 +祖细胞氧化,并通过Cox2途径用于PGD2的合成。
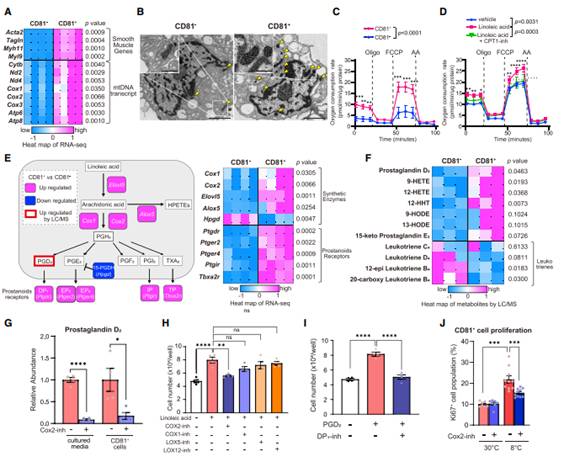
图5 CD81+在线粒体代谢和花生四烯酸途径中富集
为解决米色祖细胞如何摄取亚油酸,作者首先在腹股沟WAT中寻找CD81+米色祖细胞相对于CD81-细胞高表达的细胞膜蛋白。转录组学数据鉴定出多个候选基因,包括Slc7a2(或称为Cat2)、Tmem37和Cd36(图6A)。
作者用qPCR(图6B)和CD36抗体(图6C)在独立样本中验证CD36与CD81、整合素b1或b2和SRC家族激酶类形成异源复合物,驱动CD36的内化。。接着,作者通过选择性敲除CD81+细胞(Cd81-CreERT2Cd36flox / flox小鼠,Cd36CD81 KO小鼠)中的CD36来验证CD36介导米色祖细胞摄取亚油酸的假说。Cd36CD81 KO小鼠和同窝对照在30℃下用他莫昔芬处理,然后逐渐适应8℃3天(图6D-E)。冷暴露3天后,发现Cd36CD81 KO小鼠腹股沟WAT中Ki67+ CD81 +细胞数量显著低于对照小鼠45 %(图6F-G)。组织学分析显示,与对照小鼠相比,Cd36CD81 KO小鼠腹股沟WAT中含有更少的多房UCP1 +米色脂肪细胞(图6H)。然而,由于Cd36CD81 KO小鼠腹股沟WAT中棕色/米色脂肪选择性基因的表达水平低于对照组(图6I),因此CD36缺失导致的米色脂肪生物合成受损是显著的。
在没有任何刺激的情况下,对照组和Cd36CD81 KO来源的CD81+细胞的增殖没有差异。然而,亚油酸诱导的Cd36CD81 KO小鼠CD81+细胞增殖明显减弱(图6J)。补充亚油酸后,Cd36CD81 KO小鼠腹股沟WAT中Ki67 + CD81 +细胞数量低于对照组小鼠(图6K)。在Cd36CD81 KO小鼠中,CD81 +细胞增殖减弱导致米色脂肪生物合成受损,因为冷和亚油酸补充对米色脂肪生物合成的加和作用被削弱(图6L)。此外,与对照组小鼠相比,Cd36CD81KO小鼠腹股沟WAT部位含有较少的多房米色脂肪细胞(图6M)。最后得出结论,病理生理棕色化刺激、冷习服、b3 - AR激活和烧伤促进米色脂肪新生(图7)。
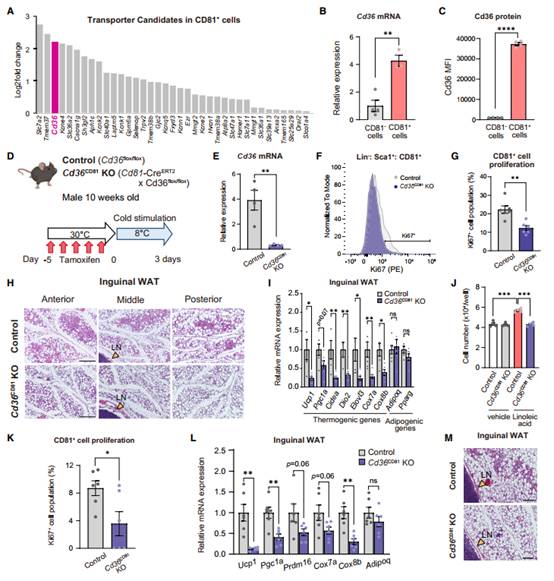
图6 CD36是CD81+ APC增殖所必需的
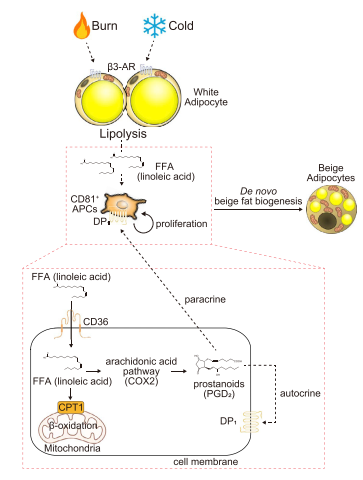
图7米色脂肪生物生成的模型
参考文献:
Abe I, Oguri Y, Verkerke ARP, Monteiro LB, Knuth CM, Auger C, Qiu Y, Westcott GP, Cinti S, Shinoda K, Jeschke MG, Kajimura S. (2022). Lipolysis-derived linoleic acid drives beige fat progenitor cell proliferation. Dev Cell;57(23):2623-2637.e8. doi: 10.1016/j.devcel.2022.11.007.