






SNORD88C促进非小细胞肺癌的生长和转移
核仁小RNA (snoRNAs)已被证明在癌症发展中发挥关键的调节作用。通过数据库和snoRNA测序筛选到长度为97 nt的SNORD88C。我们首次证实这种snoRNA在组织和血浆中上调,并可作为一种非侵入性诊断生物标志物。在体外和体内实验中证实SNORD88C促进NSCLC的增殖和转移。在机制上,SNORD88C促进了28S rRNA上C3680位点的2’- O-甲基化修饰,进而增强了下游SCD1的翻译,为SNORD88C在NSCLC中的调控提供了新的见解。本文于2022年11月发表于“Cell Death & Differentiation”(IF=12.067)上。
技术路线

结果
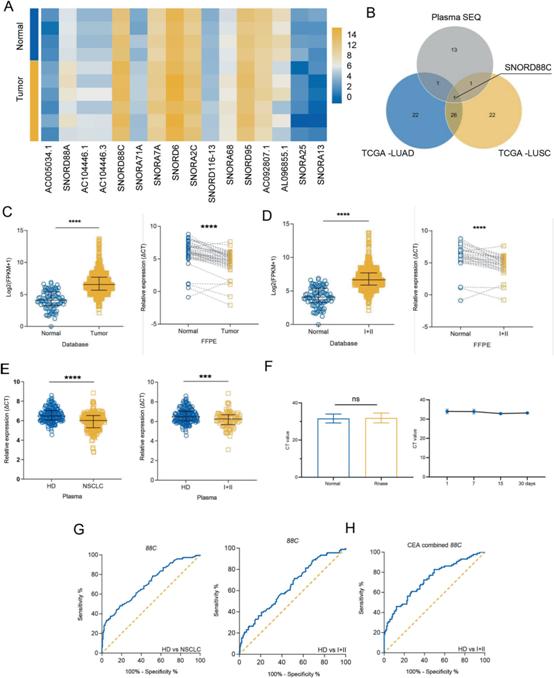
1)SNORD88C在NSCLC中上调,并作为一种非侵入性诊断生物标志物
为了识别NSCLC中异常表达的snoRNA,我们分析了TCGA SNORic数据库中肿瘤组织(包括肺腺癌(LUAD)和肺鳞癌(LUSC))和癌旁组织中snoRNA的表达差异(图S1A, B)。接下来,对4名健康供体和6名NSCLC患者的血浆进行snoRNA测序,筛选出了12个上调snoRNA和4个下调snoRNA(图1A)。最后对上述差异snoRNA进行VENN分析(图1B),只有SNORD88C重叠,被选为研究对象。我们进一步验证了SNORD88C在NSCLC中的表达上调。如图1C所示,无论是TCGA数据还是FFPE检测,肿瘤组织中SNORD88C均明显升高。TCGA数据和FFPE检测结果均显示,早期NSCLC (I + II)中SNORD88C表达较正常组织上调(图1D),提示其在NSCLC的发生发展中发挥作用。进一步验证178例健康捐赠者和224例NSCLC患者血浆中SNORD88C水平,发现其在NSCLC和早期NSCLC中上调(图1E)。SNORD88C主要在无囊泡的血浆中表达,并且在血浆中稳定(图1F)。同时对诊断效果进行ROC曲线分析,NSCLC和早期NSCLC的曲线下面积(AUC)分别为0.7097和0.6406(图1G)。癌胚抗原(CEA)是临床上公认的传统生物标志物,但对早期NSCLC的临床诊断效率较低。然而,联合SNORD88C时,早期NSCLC CEA的AUC从0.6650显著升高至0.7279(图1H)。综上所述,SNORD88C在NSCLC中上调,并作为一种非侵入性诊断生物标志物。
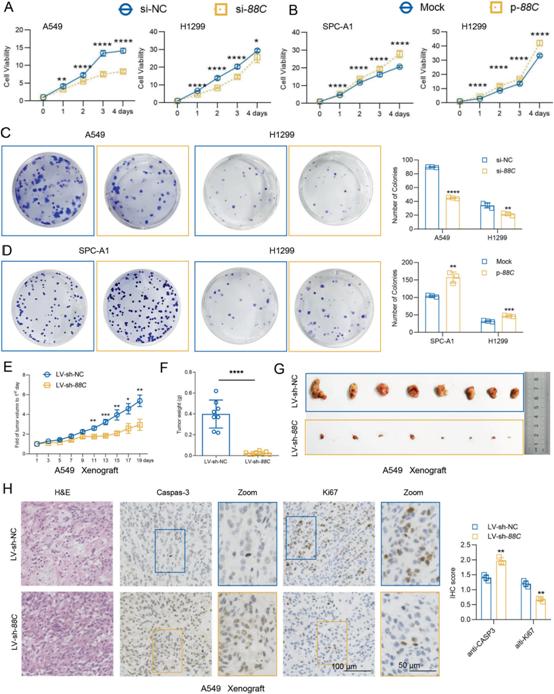
2)在体外和体内,SNORD88C促进NSCLC增殖
为了阐明SNORD88C在NSCLC进展中的作用,我们在三种NSCLC细胞系(A549、H1299和SPC-A1)中过表达或下调SNORD88C。通过CCK-8和集落形成分析来确定SNORD88C对细胞增殖的影响。如图2A所示,敲低SNORD88C显著抑制A549和H1299细胞的细胞活力;相反,强制表达SNORD88C可促进SPC-A1和H1299细胞的生长(图2B)。一致地,沉默SNORD88C的A549和H1299细胞集落形成暂停,而过表达SNORD88C导致SPC-A1和H1299细胞集落形成能力明显增强(图2C, D)。为了确定SNORD88C在体内对NSCLC增殖的影响,我们使用SNORD88C稳定敲除(LV-sh-SNORD88C)或对照(LV-sh-NC)的A549细胞建立异种移植裸鼠模型。正如预期的那样,与对照组相比,LV-sh-SNORD88C组的异种移植生长明显放缓(图2E),肿瘤重量和体积明显减小(图2F, G)。Caspase-3和Ki67免疫组化染色显示,LV-sh-SNORD88C促进A549异种移植细胞凋亡,但抑制增殖(图2H)。综上所述,在体外和体内NSCLC中,SNORD88C是一种增殖调节剂。
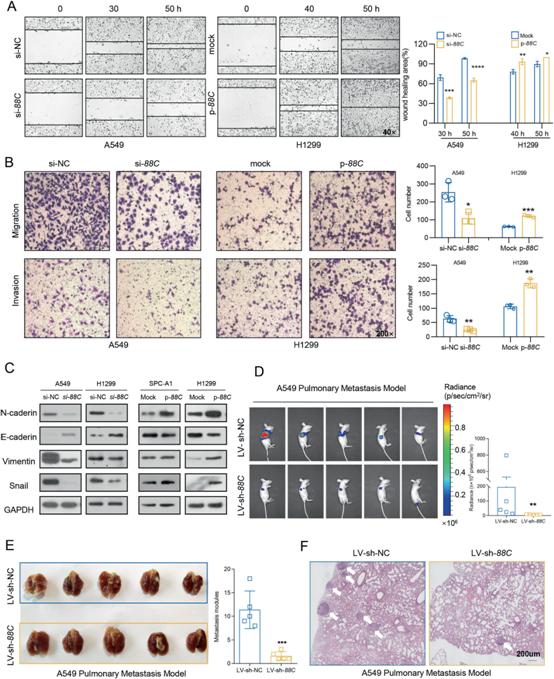
3)SNORD88C促进NSCLC在体外和体内的迁移和侵袭
我们接下来确定SNORD88C在迁移和侵袭中的作用。伤口愈合实验显示,在A549和H1299细胞中,SNORD88C的下调显著抑制了其迁移能力,而在H1299和SPC-A1细胞中,过表达则促进了其迁移能力(图3A)。此外,沉默SNORD88C表达后,A549和H1299细胞的迁移和侵袭能力明显下降,但过表达SNORD88C显著刺激了这些能力(图3B)。考虑到EMT在侵袭和迁移中的重要作用,我们检测了SNORD88C对EMT相关蛋白表达的影响。如图3C所示,在A549和H1299细胞中,敲低SNORD88C会增加E-cadherin,抑制N-cadherin, Vimentin和Snail,而在SPC-A1和H1299细胞中,过表达SNORD88C则相反。体内生物发光成像(BLI)显示LV-sh-SNORD88C组荧光素酶信号强度显著低于对照组(图3D)。与对照组相比,LV-sh-SNORD88C组小鼠肺部发生的转移位点更少(图3E)。此外,H&E染色显示LV-sh-NC组有部分转移位点侵犯肺包膜,而LV-sh-SNORD88C组未观察到(图3F)。这些结果表明,SNORD88C促进了NSCLC在体外和体内的迁移和侵袭。
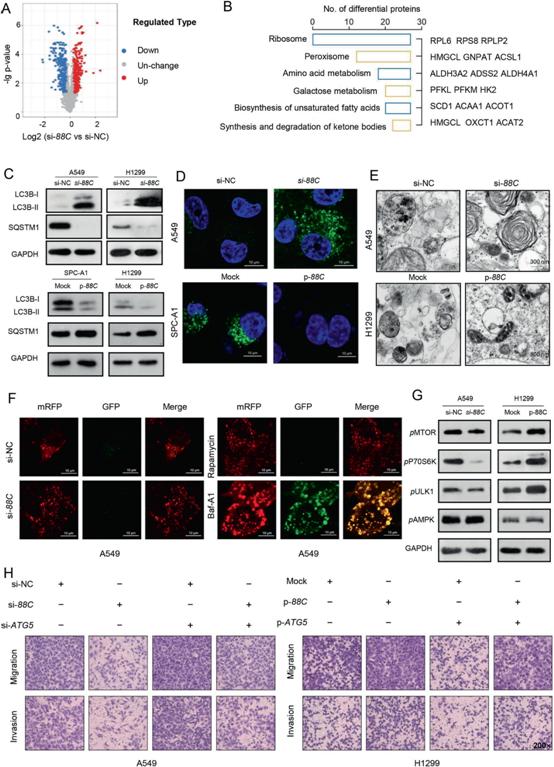
4)SNORD88C减弱的自噬是促进其迁移和侵袭所必需的
为了探索SNORD88C的下游靶基因,采用LC-MS/ MS为基础的TMT标记定量蛋白质组学方法对沉默SNORD88C或阴性对照的H1299进行研究。如图4A所示,共出现384个上调蛋白和369个下调蛋白,其中大部分富集在代谢相关通路(图4B)。由于自噬允许细胞在营养应激条件下保持代谢充足并存活,并且可以通过多种代谢途径进行调节。因此,我们进一步检测了SNORD88C对自噬的影响。如图4C所示,在A549和H1299细胞中,敲低SNORD88C均能激活自噬,而过表达SNORD88C则对SPC-A1和H1299细胞的自噬有抑制作用。相应地,免疫荧光也验证了SNORD88C敲低后自噬体数量增加,而SNORD88C过表达后自噬体数量减少(图4D)。接下来,透射电子显微镜(TEM)进一步证实了这一现象。超微结构上,敲低SNORD88C显著增加,过表达SNORD88C减少自噬液泡数量(图4E)。在SNORD88C沉默或过表达的细胞中,与雷帕霉素处理的细胞一样,大部分斑点失去了GFP信号,保留了mRFP信号,但与Baf-A1处理的细胞不同,GFP的猝灭明显减弱(图4F),说明SNORD88C对自噬通量影响不大。接下来,我们研究了mTOR/ULK1信号通路是否有助于SNORD88C减弱的自噬。如图4G所示,敲低SNORD88C导致A549中MTOR及其下游蛋白P70S6K和ULK1的磷酸化下调,而在H1299中过表达SNORD88C后,这些蛋白的磷酸化上调。值得注意的是,AMPK磷酸化不受SNORD88C的影响,这表明SNORD88C调控mTOR/ULK1信号不依赖于AMPK(图4G)。为了提供SNORD88C减弱的自噬与其诱导的肺癌细胞迁移和侵袭之间的直接联系,用ATG5 siRNA (si-ATG5)和质粒(p-ATG5)转染NSCLC细胞(图4H),同时用3-甲基腺嘌呤(3-MA)或雷帕霉素处理,然后进行迁移和侵袭检测。si-ATG5和3-MA可消除SNORD88C沉默对A549细胞迁移和侵袭的抑制作用,而过表达SNORD88C增强的H1299细胞转移作用可通过添加p-ATG5或雷帕霉素部分逆转。总之,这些结果支持SNORD88C抑制自噬,进而促进NSCLC细胞的迁移和侵袭。
5)SNORD88C通过上调SCD1蛋白表达抑制自噬
由于上述LC-MS/MS为基础TMT标记的定量蛋白质组学富集了不饱和脂肪酸生物合成通路,因此我们对SNORD88C敲除的H1299细胞进行了以脂肪酸为目标的GC-MS代谢组学研究,以确定SNORD88C减弱自噬的代谢产物。如图5A所示,与对照组相比,SNORD88C敲除的细胞中多种不饱和脂肪酸均有降低,其中单不饱和脂肪酸(MUFA)油酸(OA)的下降更为明显,说明SNORD88C在脂质积累和过氧化过程中起着至关重要的作用。因此,采用C11-bodipy染色,流式细胞术分析显示沉默SNORD88C促进脂质过氧化,过表达SNORD88C抑制脂质过氧化(图5B)。
SCD1是合成MUFA的中心脂肪生成酶,在沉默SNORD88C后,其在上述LC-定量蛋白质组学中也表现出最显著的变化。重要的是,western blot检测成功验证了SNORD88C正调控SCD1蛋白水平,在A549和H1299细胞中,SNORD88C下调后SCD1蛋白水平下降,但在H1299和SPC-A1细胞中,过表达SNORD88C后SCD1蛋白水平显著升高(图5C)。流式细胞仪分析显示,SCD1的过表达和敲低分别能成功逆转沉默SNORD88C以及过表达SNORD88C对脂质过氧化的促进和抑制作用(图5D),提示SNORD88C通过上调SCD1蛋白表达抑制脂质过氧化。此外,我们还探讨了SCD1在SNORD88C减弱的自噬中的作用。如图5E所示,过表达SCD1可以通过下调SNORD88C恢复A549细胞的自噬激活。一致地,SCD1沉默缓解了SNORD88C过表达对自噬的抑制。此外,我们还观察了SNORD88C和SCD1对mTOR通路的影响。正如预期的那样,SCD1消除了SNORD88C对mTOR通路相关蛋白的作用,包括pMTOR、pP70S6K、pULK1(图5F)。最后,我们研究了SNORD88C/SCD1轴与肺癌细胞迁移和侵袭之间的直接联系。如图5G和5H所示,SCD1过表达逆转了SNORD88C沉默对A549和H1299细胞迁移和侵袭的抑制作用,而SCD1下调则取消了SPC-A1和H1299细胞中SNORD88C过表达对肺癌细胞转移的促进作用。综上所述,SCD1是SNORD88C的下游调控蛋白,通过抑制自噬促进NSCLC细胞的迁移和侵袭。
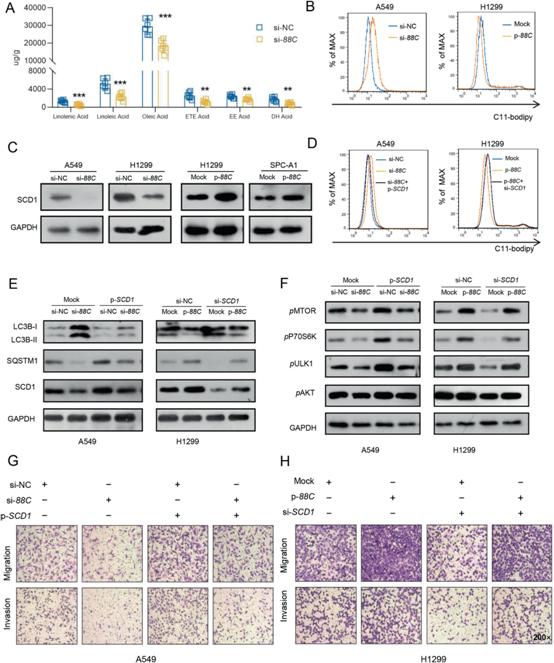
6)SNORD88C通过引导28S rRNA的2’-O-me调控SCD1的翻译活性
为了探索SNORD88C/ SCD1轴相关的潜在机制,我们首先检测了SNORD88C对SREBP1的影响,SREBP1是调控SCD1表达的重要转录因子。出乎意料的是,SNORD88C对SREBP1的表达没有明显的影响(图6A),说明SNORD88C没有调节SCD1的转录,这也被另一个观察结果证实,即SNORD88C不影响SCD1的mRNA水平(图6B)。由此,我们推测SNORD88C在转录后水平调控SCD1的表达。接下来,使用蛋白酶体抑制剂MG132阻断蛋白酶体降解。然而,MG132处理明显增加了SCD1蛋白水平,但未能逆转SNORD88C对SCD1的调控(图6C),这表明涉及一种蛋白降解无关的方式。
先前的研究表明,box C/D snoRNAs在rRNA生物发生中起着重要作用,因为它们参与核仁中2 '-O-me的引导,进而调节细胞质中蛋白质的翻译。首先,我们使用cy3标记的探针靶向SNORD88C进行荧光原位杂交(FISH)检测细胞分布。如图6D所示,SNORD88C主要位于细胞核和细胞质中。通过细胞质/核分离的q-PCR进一步验证了这一结果(图6E),表明SNORD88C具有直接调节rRNA生物学特性的潜力。此外,我们基于生物信息学通过snoPY数据库预测了SNORD88C靶向的rRNA,发现只有28S rRNA含有与SNORD88C高度互补的序列,更重要的是c3680的2 '-O-me的潜在位点(图6F)。然后,使用能够区分前体(ITS-28S)和总28S rRNA的引物进行量化,然而,无论是在敲除SNORD88C的A549细胞中,还是在过表达SNORD88C的H1299细胞中,成熟28S rRNA均未观察到明显变化(图6G)。接下来,采用RTL-P实验确认SNORD88C引导的28S rRNA的2’-O-me。在A549细胞中,低浓度dNTPs时,敲低SNORD88C可显著降低C3680位点28S rRNA的2'-O-me活性,而高浓度dNTPs时则不会(图6H)。同时,过表达SNORD88C可提高低浓度dNTPs时2'-O-me活性(图6I)。已有研究表明2'-O-me可提高rRNA诱导的翻译准确性,进而延长蛋白质半衰期。因此,通过添加环己亚胺(CHX)来进行体外翻译试验。当用CHX处理SNORD88C敲除的A549细胞时,SCD1蛋白的半衰期缩短(图6J),而在过表达SNORD88C的H1299细胞中,SCD1蛋白的半衰期延长(图6K)。最后,为了确定SNORD88C直接影响翻译的可能性,对其进行了多核糖体分析。如图6L, M所示,SNORD88C沉默或过表达不影响核糖体部分的分布轮廓,表明SNORD88C不影响全局翻译。然而,SNORD88C沉默显著降低了SCD1 mRNA在单体和多体部分的富集,使其分布从较重的核糖体部分向较轻的核糖体部分分布,而SNORD88C过表达则相反,更直接地证明了SNORD88C通过影响翻译水平调控SCD1的表达。
结论:
我们发现SNORD88C是NSCLC中一种新的致癌snoRNA。我们证明了SNORD88C在组织和血浆中上调,并可作为一种非侵入性诊断生物标志物;在体外和体内均证实其促进NSCLC的增殖和转移。重要的是,SNORD88C引导28S rRNA甲基化,显著提高下游靶基因SCD1的翻译活性。SCD1作为MUFA合成的限速酶,抑制自噬进一步促进转移,为SNORD88C在NSCLC中的调控提供了新的视角。
参考文献:
Wang K, Wang S, Zhang Y, Xie L, Song X, Song X. SNORD88C guided 2'-O-methylation of 28S rRNA regulates SCD1 translation to inhibit autophagy and promote growth and metastasis in non-small cell lung cancer. Cell Death Differ. 2022 Nov 14. doi: 10.1038/s41418-022-01087-9.