






LncRNA CEBPA-DT通过与hnRNPC相互作用激活DDR2/β-catenin从而促进肝癌转移
原发性肝癌(PLC)是第六大最常见的人类癌症类型,在全球癌症死亡率中排名第三。肝细胞癌(HCC)是肝癌的主要组织学类型,约占PLC的90%。尽管在检测和治疗方面不断改善,但由于肝细胞癌患者进展迅速,术后肿瘤复发和转移的患病率高,HCC患者的远期结局仍然较差,5年总生存率低于30%。因此,阐明HCC生长和进展的分子机制以确定新的治疗靶点至关重要。长非编码RNA CEBPA-DT是CCAAT增强子结合蛋白α(CEBPA)基因的不同转录本,已被证明参与多种肿瘤进展。然而,还没有研究在HCC中建立其促癌机制。该研究发表于《Journal of Experimental & Clinical Cancer Research》,IF: 12.658。
技术路线:
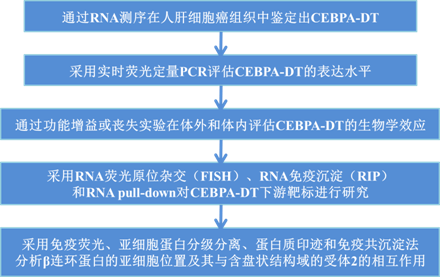
主要研究结果:
1. CEBPA-DT在肝细胞癌组织中上调,与患者预后相关
为了筛选与肝细胞癌转移相关的lncRNA,采用RNA-seq分析术后肝外转移患者的11个HCC组织和未发生转移或复发的患者的11个HCC组织之间的差异表达基因,鉴定出320个显著改变的lncRNA(图1A)。6个lncRNA(CEBPA-DT,编号:AL136372,MEG3,DBH-AS1,TUG1,PSMA3-AS1)在肝外转移患者的11个HCC组织中具有最高的丰度(平均FPKM > 5)。CEBPA-DT被选为进一步研究的候选lncRNA,因为它是在前面提到的6个候选lncRNA中(图1B,C)最显著的上调靶标(log2(倍数变化)= 5.753,P = 0.013)。此外,研究者检查了CEBPA-DT在54个具有相应正常肝组织的肝细胞组织中的表达水平。RT-qPCR分析表明,肝癌组织中CEBPA-DT的表达显著高于配对正常肝组织中的表达(图1D)。在TCGA LIHC数据中的50对肝细胞癌和正常组织中也观察到了类似的结果(图1F)。
为了进一步评估HCC患者中CEBPA-DT表达水平的预后价值,在88个HCC组织中测量了CEBPA-DT的表达水平。患者分为高CEBPA-DT组(n=44)和低CEBPA-DT组(n=44),符合CEBPA-DT在HCC组织中的表达。较高的CEBPA-DT表达水平与血清甲胎蛋白(AFP)水平升高、肿瘤体积较大、肿瘤分化差和微血管浸润(MVI)显著相关,提示CEBPA-DT表达较高与肝细胞癌生长和转移有关。Kaplan-Meier生存曲线表明,高CEBPA-DT水平组的HCC患者与低水平CEBPA-DT患者的总生存期(OS)和无复发生存期(RFS)显著相关(图1E)。一致地,从TCGA LIHC队列中获得相同的结果(图1G)。在单因素分析中,HBsAg阳性状态,肿瘤分化差,肿瘤大小>5cm,存在MVI,AFP>400ng/μL,远处转移和高CEBPA-DT表达被确定为与HCC患者OS和RFS阴暗相关的潜在危险因素。这些变量进一步包含在多元回归模型中。结果显示,MVI和高CEBPA-DT表达是OS和RFS的独立危险因素。
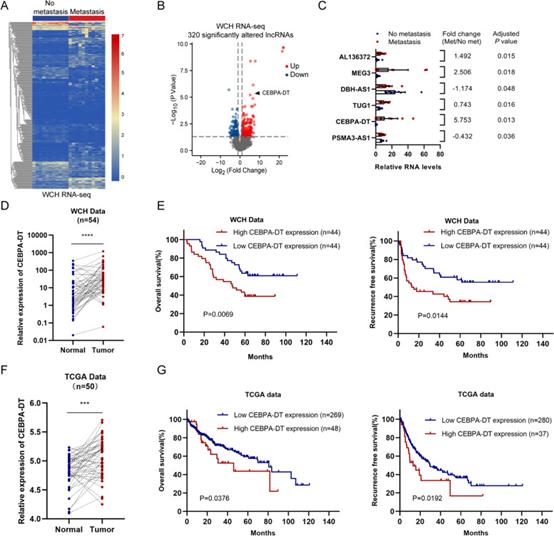
图1 CEBPA-DT 在伴有肝外转移的肝细胞癌中上调,并与患者预后相关
2. CEBPA-DT促进肝癌细胞在体外和体内的增殖和转移
选取7个人肝癌细胞系,采用RT-qPCR检测CEBPA-DT的表达水平,结果表明,HCCLM9、SNU-449和SK-Hep1的CEBPA-DT表达水平高于Huh7、Hep3B和PLC/PRF/5。体外功能研究表明,沉默CEBPA-DT显著抑制HCCLM9和SNU-449细胞的细胞增殖(图2A,B)。同时,跨孔迁移、侵袭和伤口愈合迁移试验表明,肝癌细胞中CEBPA-DT下调后,细胞运动受到显著抑制(图2C-F)。此外,采用流式细胞术研究了细胞凋亡和细胞周期分布,结果表明,CEBPA-DT的耗竭显著提高了肝癌细胞的凋亡速率(图2G,H)。同时,在沉默CEBPA-DT后,观察到G0/G1期肝癌细胞百分比增加,S期肝癌细胞百分比降低(图2I,J)。接下来,通过Huh7,Hep3B和SK-Hep1细胞中的慢病毒感染构建过表达CEBPA-DT的稳定肝癌细胞系。CEBPA-DT的过表达显著提高了Hep3B和Huh7细胞的生长能力(图3A,B)。在过表达肝癌细胞的CEBPA-DT中观察到细胞运动的显著增强(图3C-F)。细胞凋亡试验表明,CEBPA-DT过表达降低了Huh7和Hep3B细胞的凋亡率(图3G,H)。细胞周期分析显示,CEBPA-DT过表达增加了S级分,降低了G0/G1级分(图3I,J)。
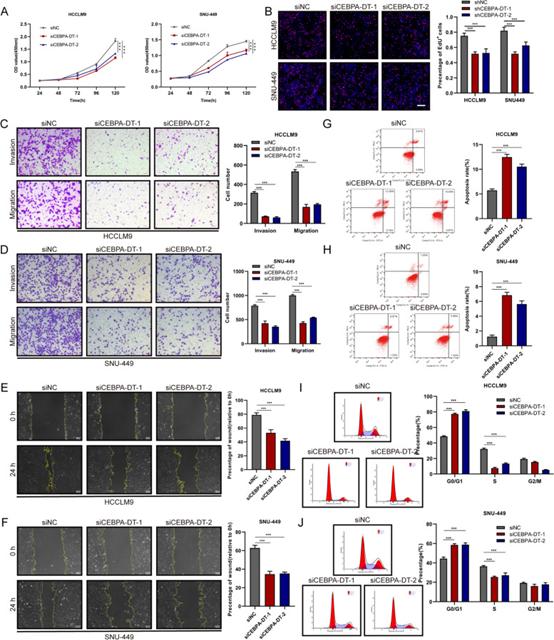
图2 CEBPA-DT的沉默在体外抑制肝癌细胞的肿瘤生长和转移
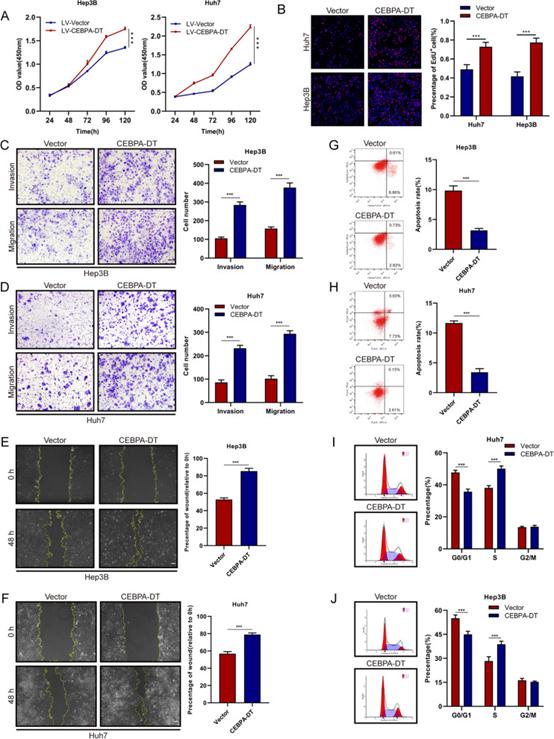
图3 CEBPA-DT在体外促进肝癌细胞的肿瘤生长和转移
研究者进一步确定了CEBPA-DT对体内致瘤性的影响。建立CEBPA-DT沉默HCCLM9细胞和CEBPA-DT过表达SK-Hep1细胞,并皮下注射到裸鼠体内。当CEBPA-DT被敲低时,观察到肿瘤体积和重量显著降低,当CEBPA-DT过表达时,获得相反的结果(图4A,B),表明CEBPA-DT促进体内肿瘤生长。为进一步评价CEBPA-DT对体内肿瘤转移的影响,建立了肝原位植入模型和肺转移模型。6周后,在肝脏中,CEBPA-DT沉默组观察到的GFP荧光强度低于对照组(图4C),当CEBPA-DT过表达时表现出相反的结果(图4D)。IVIS拍摄结果表明,CEBPA-DT敲低组与比对照组更弱的肺部生物发光信号强度相关(图4G);同时,与空载体组相比,CEBPA-DT过表达增强了肺部的生物发光活性(图4H)。苏木精和伊红(HE)染色结果显示,CEBPA-DT沉默组的转移灶数量均在肝脏中减少(图4E)。相比之下,与对照组相比,CEBPA-DT过表达组的肝肺转移灶数量增加(图4F,J)。综上所述,CEBPA-DT是一种促癌lncRNA,可促进肝癌细胞在体外和体内的增殖和转移。
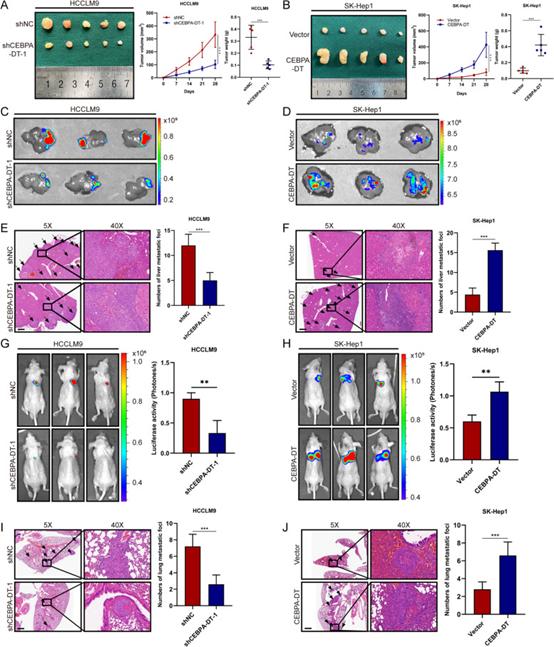
图4 CEBPA-DT促进体内肿瘤生长和转移
3. CEBPA-DT与RNA结合蛋白hnRNPC相互作用
为了验证CEBPA-DT是否充当“miRNA海绵”,研究者进行了生物信息学分析,以使用Starbase评估CEBPA-DT的潜在结合miRNA。结果表明,has-miR-532-3p是最具约束力的靶标。研究者接下来建立了CEBPA-DT野生型和突变荧光素酶质粒;通过野生型/突变的CEBPA-DT与NC模拟物/ miR-532-3p的共转染,通过Rluc/Fluc测量相对荧光素酶活性。为了验证CEBPA-DT是否通过与蛋白质结合来调节肿瘤生长和转移,研究者接下来进行了生物素标记的RNA pull-down和质谱(MS)以筛选与肝癌细胞中CEBPA-DT相互作用的蛋白质。由于RNA FISH和先前进行的亚细胞RNA分级分离测定证明CEBPA-DT主要位于细胞核中(图5A),仅使用核提取物进行RNA pull-down测定。银染在近40 kD处鉴定出特定的蛋白质条带,随后通过MS检测(图5B)。共鉴定出177种蛋白质作为CEBPA-DT相互作用靶标。其中,RNA结合蛋白异质核糖核蛋白C(hnRNPC)得分最高(图5B)。
为了验证CEBPA-DT与hnRNPC之间的相互作用,研究者确定了hnRNPC的亚细胞位置。接下来,生物素标记的RNA pull-down和RIP测定进一步证实了CEBPA-DT可以与hnRNPC相互作用(图5C,D)。为了进一步验证CEBPA-DT和hnRNPC之间的特异性相互作用位点,研究者构建了三个CEBPA-DT截短片段(1-750nt,751-1500nt和1501-2252nt)(图5E),以及基于RNA识别基序(RRM,16-87aa)的截短hnRNPC片段(图5G)。RNA pull-down测定显示,751-1500个核苷酸负责CEBPA-DT与hnRNPC的RNA识别基序之间的相互作用(图5F,H)。先前的一项研究证明,hnRNPC通过与c-myc相互作用,从细胞核部分地重新定位到细胞质。研究者假设CEBPA-DT和hnRNPC之间的相互作用可以激活hnRNPC的亚细胞重定位。亚细胞蛋白分离测定显示,CEBPA-DT的过表达上调了细胞质中hnRNPC的蛋白水平,同时降低了细胞核中的hnRNPC水平(图5I)。通过RNA FISH结合IF测定获得了类似的观察结果(图5J)。综上所述,这些数据表明CEBPA-DT可能在物理上结合hnRNPC并诱导hnRNPC从肝癌细胞的细胞核到细胞质的转定位。
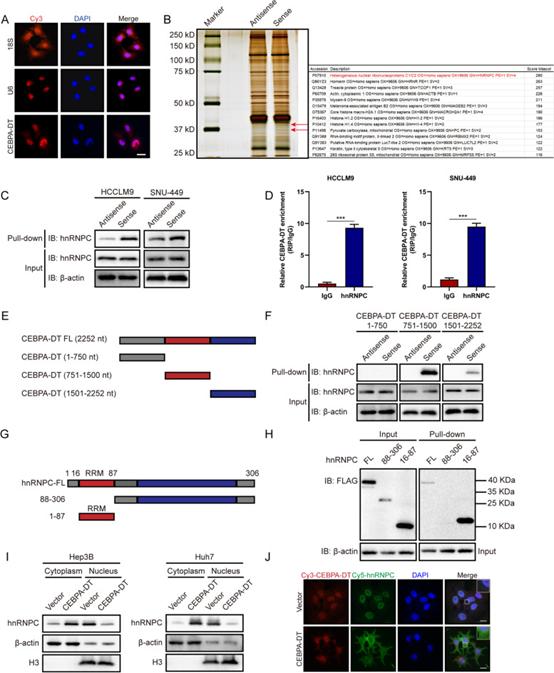
图5 CEBPA-DT与RNA结合蛋白hnRNPC相互作用
4. CEBPA-DT通过hnRNPC-DDR2轴促进肝癌细胞的转移
更深入地了解CEBPA-DT在肝癌细胞中的致瘤作用。进行RNA测序以分析受CEBPA-DT沉默影响的转录组变化。聚类分析显示,CEBPA-DT沉默HCCLM9细胞中共有2040个下调基因。不同数据库中的进一步富集分析表明,细胞外基质相关途径和基因表现出最显著的改变(图6A)。这些分析的交集揭示了8个下调基因,这些基因受到RT-qPCR的影响,结果表明,沉默CEBPA-DT后,DDR2的mRNA水平下调最显著(图6B)。
作为普遍表达的异质核糖核蛋白(hnRNPs)家族的成员,hnRNPC作为RBP结合核RNA,对于许多mRNA的稳定和翻译至关重要,通常与靶mRNA的3' UTR区域相互作用。考虑到CEBPA-DT诱导hnRNPC从细胞核向细胞质的重新定位,以及CEBPA-DT调控DDR2 mRNA水平,研究者假设CEBPA-DT可能增强DDR2 mRNA的稳定性,从而通过增加hnRNPC的细胞质表达来促进翻译。为了验证这一猜想,研究者测量了用siRNA沉默hnRNPC后DDR2的mRNA和蛋白质水平。结果表明,DDR2 mRNA和蛋白质水平显著下调(图6C,D)。更关键的是,在CEBPA-DT上调后,hnRNPC与DDR2 mRNA之间的相互作用明显增强(图6E)。总之,这些结果表明DDR2是CEBPA-DT-hnRNPC复合物的下游靶标。
研究者相继研究了CEBPA-DT-hnRNPC轴对DDR2下调的影响机制。通过放线菌素D处理肝癌细胞,研究者发现CEBPA-DT的过表达降低了DDR2 mRNA的降解,延长了其半衰期,从而增加了DDR2 mRNA的表达,同时hnRNPC的缺失显著抑制了这种促进,提高了DDR2 mRNA水平(图6G)。一致地,在CEBPA-DT上调和hnRNPC沉默后,蛋白质水平表现出相同的变化趋势(图6F)。
先前的一项研究阐明DDR2有助于HCC侵袭和转移。研究者的体外功能研究表明,在沉默DDR2的mRNA和蛋白质水平后,肝癌细胞的运动性受到显著抑制。在功能上,伤口愈合和Transwell测定表明,CEBPA-DT的过表达可以增强肝癌细胞的运动性,而DDR2的缺失可以阻止这种增强。裸鼠肺转移模型也获得了类似的结果(图6H)。此外,通过在50个HCC样品中应用IHC和RT-qPCR,研究者发现与正常组织相比,HCC组织中DDR2的蛋白质和mRNA水平明显增加(图6I)。此外,DDR2的表达与HCC组织中CEBPA-DT的表达呈正相关(图6J)。综上所述,上述数据表明CEBPA-DT可以通过hnRNPC-DDR2轴促进HCC的转移。
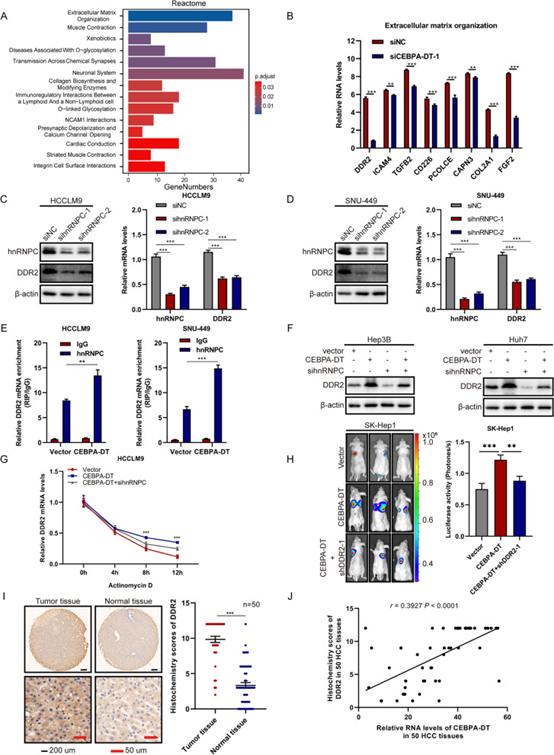
图6 CEBPA-DT通过hnRNPC-DDR2轴促进肝癌细胞的转移
5. CEBPA-DT诱导肝癌细胞中的EMT
先前的研究表明,CEBPA-DT下调改变了EMT相关基因的表达;然而,CEBPA-DT参与EMT过程的分子机制尚不清楚。本研究首先采用RT-qPCR筛选了几种EMT标志物,包括上皮标志物E-钙粘蛋白(CDH1)、间充质标志物N-钙粘蛋白(CDH2)、Vimentin(VIM)、SNAIL1、SNAIL2、ZEB1、TWIST1和FOXC1,结果表明,敲低CEBPA-DT诱导上皮标志物CDH1的表达,同时抑制间充质标志物(CDH2、VIM、SNAIL1)的表达。同时,CEBPA-DT的过表达表现出相反的结果。在ZEB1,SNAIL2,TWIST1和FOXC1中没有观察到显著的变化(图7A)。为了检查EMT转录因子Snail1是否被CEBPA-DT激活,将Snail1启动子荧光素酶报告质粒转染到肝癌细胞中。结果表明,CEBPA-DT沉默显著降低了Snail1启动子的活性,而上调的CEBPA-DT显著诱导了启动子活性(图7B)。蛋白质印迹实验证实,CEBPA-DT过表达抑制了E-钙粘蛋白的蛋白表达,诱导了肝癌细胞中N-钙粘蛋白、Vimentin、Snail和DDR2的表达,而CEBPA-DT下调的结果相反(图7C)。根据蛋白质印迹测定,IF实验表明,CEBPA-DT的消耗显著增加了E-钙粘蛋白的表达,并降低了Vimentin和Snail的表达(图7D)。为了进一步验证EMT过程是通过CEBPA-DT激活Snail1诱导的,研究者在Huh7细胞中敲低了Snail1。CEBPA-DT上调促进间充质标志物的表达因沉默蜗牛1而减弱;在上皮标志物E-钙粘蛋白的表达中观察到相反的趋势(图7E)。综上所述,这些数据表明CEBPA-DT通过Snail1的转录激活诱导肝癌细胞中的EMT。
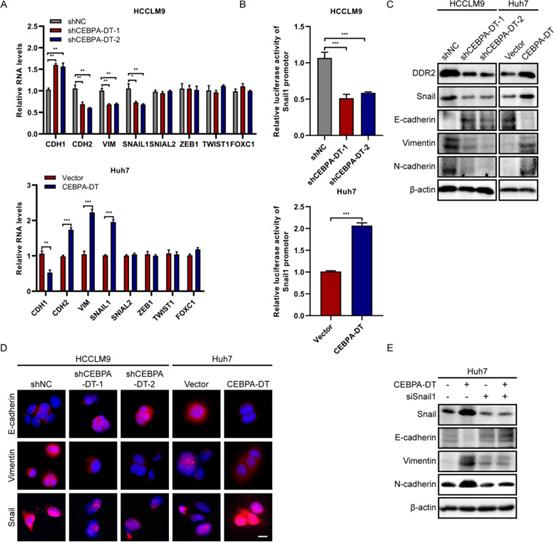
图7 CEBPA-DT在肝癌细胞中诱导EMT
6. CEBPA-DT通过DDR2和β-连环蛋白之间的相互作用促进EMT过程
作为Wnt/β-连环蛋白通路的核心成分,β-连环蛋白被发现在HCC转移中被定期激活,并与EMT过程密切相关。因此,研究者假设β-连环蛋白参与了CEBPA-DT调节的EMT过程。首先,亚细胞蛋白分级分离显示,细胞核组分中的β-连环蛋白表达在CEBPA-DT过表达肝癌细胞中显著上调,在CEBPA-DT沉默细胞中下调,而细胞质部分β-连环蛋白的表达保持不变(图8A)。接下来,免疫荧光测定表明,在CEBPA-DT沉默时,核β-连环蛋白显著重新定位到肝癌细胞的细胞质中。相反,CEBPA-DT上调后促进了β连环蛋白的核易位(图8B)。为了探索Wnt/β-连环蛋白信号传导的转录活性,进行了TOP/FOP-Flash荧光素酶报告测定,结果表明,在HCCLM9载体细胞和β连环蛋白过表达细胞中敲低CEBPA-DT降低了转录活性(图8E)。
为了评价β连环蛋白对蜗牛1启动子活性的调控能力,研究者进行了生物信息学分析,预测了β-连环蛋白在蜗牛1启动子区域的结合位点(图8C)。进一步的 ChIP-qPCR 测定表明 Snial1 启动子区域的结合位点 1 上β-连环蛋白显著富集;同时,CEBPA-DT沉默可以明显抑制这种富集(图8D)。
早期的一项研究已经证明了DDR2在EMT过程中的调节作用。研究者的数据证实DDR2受CEBPA-DT监管。因此,研究者推测DDR2和β-连环蛋白之间存在相互作用,这种相互作用是CEBPA-DT调控EMT过程所必需的。首先,使用针对DDR2的特异性抗体进行免疫共沉淀测定,结果表明DDR2确实可以与β-连环蛋白相互作用,并且在CEBPA-DT沉默后相互作用减弱,并在CEBPA-DT过表达后得到加强(图8F)。接下来,使用一种特异性抑制剂DDR2-IN-1来抑制DDR2。随着DDR2-IN-1的存在,由CEBPA-DT过表达调节的β-连环蛋白的核易位增加显著耗尽(图8G)。最后,蛋白质印迹试验显示DDR2抑制可能导致CEBPA-DT诱导EMT过程的抑制;β-连环蛋白的过表达挽救了这种抑制(图8H)。综上所述,CEBPA-DT通过DDR2与β-连环蛋白的相互作用促进了EMT过程。
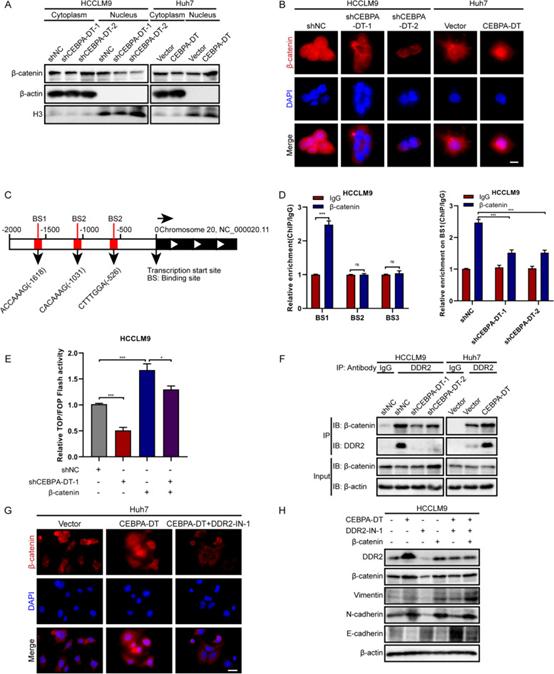
图8 CEBPA-DT通过DDR2和β-catenin之间的相互作用促进EMT过程
结论:
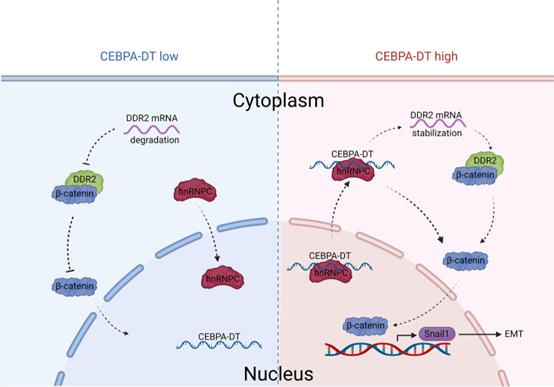
综上所述,该研究将CEBPA-DT确定为HCC转移的效应物。此外,研究者发现CEBPA-DT通过与hnPNPC相互作用激活DDR2/β-连环蛋白轴来促进HCC的进展和转移。重要的是,该研究结果为lncRNA参与HCC转移的潜在分子机制提供了新的见解,并为进一步的CEBPA-DT靶向HCC治疗方法奠定了基础。
示意图:
说明CEBPA-DT在肝癌转移中的作用
参考文献:
Cai Y, Lyu T, Li H, Liu C, Xie K, Xu L, et al. LncRNA CEBPA-DT promotes liver cancer metastasis through DDR2/β-catenin activation via interacting with hnRNPC. J Exp Clin Cancer Res. 2022 Dec 6;41(1):335. doi: 10.1186/s13046-022-02544-6.