






一种由circINSIG1编码的新蛋白通过促进结直肠癌中INSIG1的泛素依赖性降解来编码胆固醇代谢
缺氧是实体肿瘤的一个标志,并导致癌细胞的代谢重编程。在结肠直肠癌(CRC)中,表观遗传调控在缺氧和异常胆固醇代谢之间的作用尚不清楚。采用高通量RNA测序方法,在常氧和低氧培养的结直肠癌细胞中鉴定低氧反应环状RNA (circRNAs)并通过多体分析和LC-MS鉴定了circINSIG1的蛋白编码潜能。circINSIG1的功能在体外和体内通过功能增益或丧失试验得到验证并通过免疫沉淀分析得出机制结果。结果发现了一种新的低氧应答环状RNA circINSIG1,该环状RNA在结直肠癌组织中表达上调,并与晚期临床分期和低生存率相关。机制上,circINSIG1编码一个121个氨基酸的蛋白circINSIG1-121,通过募集CUL5-ASB6复合物(一种泛素E3连接酶复合物),促进赖氨酸156和158处关键胆固醇代谢调节因子INSIG1的k48连锁泛素化,从而诱导胆固醇生物合成,促进结直肠癌的增殖和转移。原位异种移植肿瘤模型和患者来源的异种移植模型进一步确定了circINSIG1在CRC进展中的作用和CRC的潜在治疗靶点。circINSIG1具有表观遗传机制,为研究缺氧与胆固醇代谢之间的串扰提供了新的思路,为CRC的治疗提供了一个有前景的靶点。本文于2023年4月发表在《Molecular Cancer》IF:41.444期刊。
技术路线:

主要实验结果:
1、CRC中缺氧诱导的circINSIG1的特征及临床意义
为了鉴定CRC中与缺氧相关的环状RNA,作者对HCT8、HCT116和DLD1细胞在常氧或缺氧(1% O2, 48 h)培养下进行了环状RNA测序,结果显示,这三种细胞系中有18个环状RNA失调(图1A)。接着,选择在缺氧条件下具有相同趋势的四个circRNA ((circNPHP4, circINSIG1, circGART 和 circZDHHC5)进一步鉴定(图1B)。通过实时荧光定量PCR (qRT-PCR)分析这些circRNAs在CoCl2诱导的细胞假性缺氧模型和32对CRC样本中的表达水平,结果显示,circINSIG1(hsa_circ_0133744)在CoCl2处理的CRC细胞系和CRC组织样本中显著上调(图1C, S1A和S1B),因此选择circINSIG1进行进一步研究。进一步分析了85个配对CRC样本中circINSIG1的表达水平,证实了circINSIG1在CRC中表达上调(图1D)。结果表明,circINSIG1在晚期T期或临床分期患者中表达更高(图1E - F)。
circINSIG1是胰岛素诱导基因1 (INSIG1)的外显子3和4与292nt反向剪接而形成的(图1G)。Sanger测序与之吻合,验证了circINSIG1的反向剪接连接(图1H)。作者采用发散引物和收敛引物分别检测互补DNA (cDNA)和基因组DNA (gDNA)中的circINSIG1和INSIG1的线性转录本,结果表明,circINSIG1只能通过不同的引物在cDNA中扩增(图1I)。半衰期分析显示circINSIG1比线性INSIG1 mRNA稳定得多(图1J)。此外,circINSIG1被观察到抵抗RNase R的消化(图1K)。核质量分离实验和荧光原位杂交(FISH)实验表明,CRC细胞的细胞质中富集了circINSIG1(图1L-M)。
作者通过原位杂交分析了大量CRC患者中circINSIG1的表达(n=227)。结果还证实,与配对的正常邻近组织相比,CRC组织中circINSIG1的表达水平升高(图1N)。队列中circINSIG1的表达与CRC患者TNM分期、临床分期等临床病理特征显著相关(p<0.05)。此外,Kaplan-Meier曲线显示,高circINSIG1表达与CRC患者较差的生存率相关(图1O)。综上所述,这些结果表明了circINSIG1的循环性和临床意义。
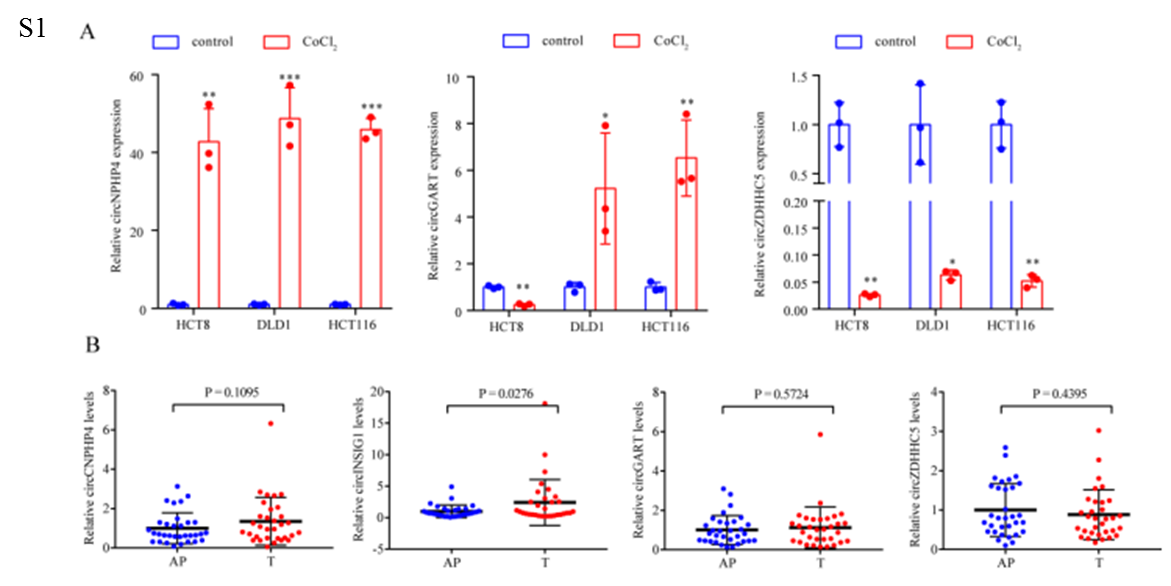
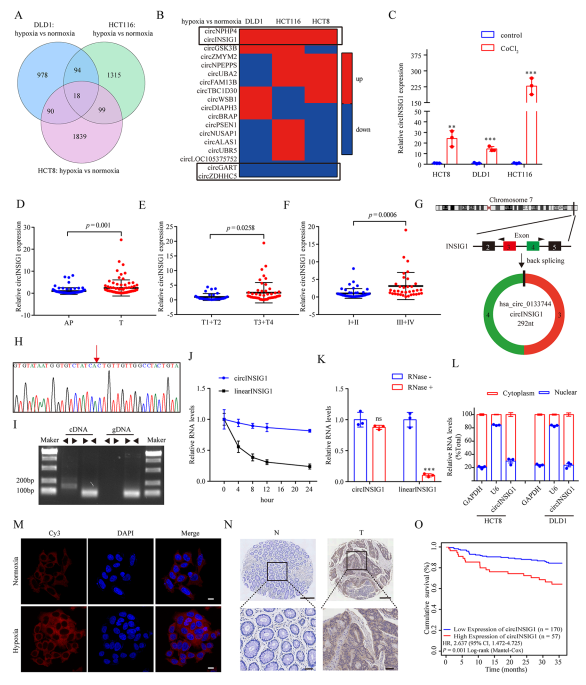
图1 CRC中缺氧诱导的circINSIG1的特征及临床意义
2、circINSIG1编码121个氨基酸的与CRC的不良预后相关的新蛋白
为了确定circINSIG1的蛋白质编码潜力,作者首先分析了circRNADb数据库中注释的circINSIG1的开放阅读框(ORF)和内部核糖体进入位点(IRES)。结果显示,circINSIG1序列中包含一个具有编码121个氨基酸蛋白潜能的跨越连接ORF(以下简称circINSIG1-121)和一个位于207-292 nt的IRES(图2A)。接着,作者进行了基于蔗糖密度梯度离心的多体分析,细胞裂解液中的核糖体分为40和60 S核糖体亚基,以及单体(80 S)和多体(图2B)。线性INSIG1 mRNA作为阳性对照,circCAMSAP1作为阴性对照,circCAMSAP1是作者在之前的研究中发现的不具有蛋白质编码能力的circRNA。、结果显示circINSIG1可以在单体和多体中检测到(图2B)。此外,通过双荧光素酶测定验证了IRES驱动ORF翻译的活性(图S2A和S2B)。由于在第一轮读取中缺少终止密码子,circINSIG1-121在第二轮翻译中由自然移码形成的C端有一条独特的20个氨基酸的尾巴(图2A)。接着制备了一种针对尾20个氨基酸序列的单克隆抗体,用于检测circINSIG1-121的表达。构建circINSIG1-flag载体,转染CRC细胞,结果显示,Flag抗体在转染circINSIG1-flag载体的细胞中检测到14KD条带,而在转染对照载体的细胞中未检测到(图2C,左)。circINSIG1-121抗体证实了circINSIG1-121的内源性存在(图2C,右)。使用过表达circINSIG1的CRC细胞进行质谱分析和SDS-PAGE进一步验证了circINSIG1-121蛋白的特异性肽片段与预测的分子量(图2D-E和S2C)。
接下来,分别通过免疫组化(IHC)和western blot分析20例和10例配对CRC样本中circINSIG1-121和INSIG1的蛋白水平。结果显示,circINSIG1-121在结直肠癌组织中表达上调,而INSIG1蛋白水平在配对的正常邻近组织和结直肠癌组织中无显著差异(图2F-G)。此外,通过免疫组化(IHC)在结直肠癌队列中检测了circINSIG1-121和INSIG1蛋白水平(n=227)。结果表明,高circINSIG1-121表达与CRC患者较差的生存率相关(图2H)。而在本研究结果中或TCGA数据库中,INSIG1的线性表达水平与CRC患者的生存率没有相关性(图2I-J)。总之,这些数据证实circINSIG1编码一种121个氨基酸的新蛋白,该蛋白与CRC的不良预后相关。
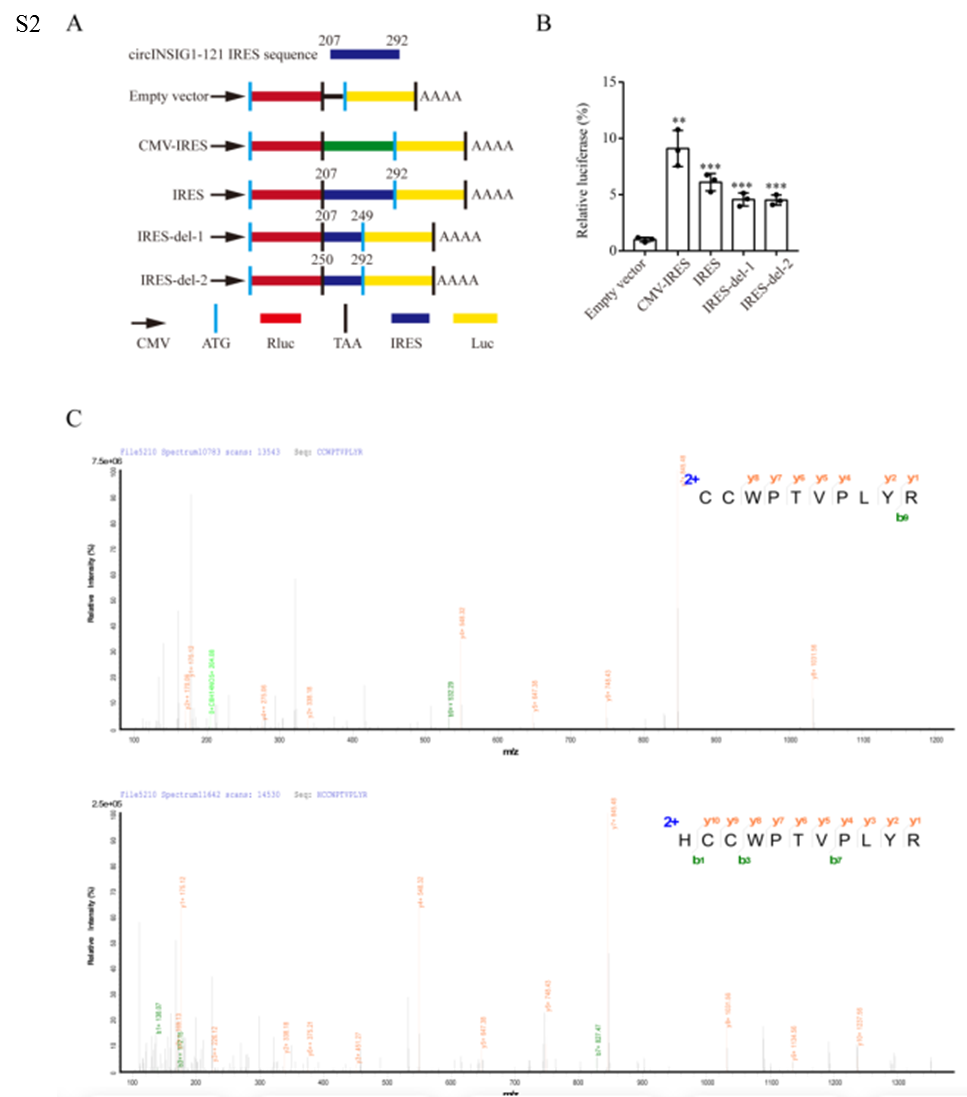
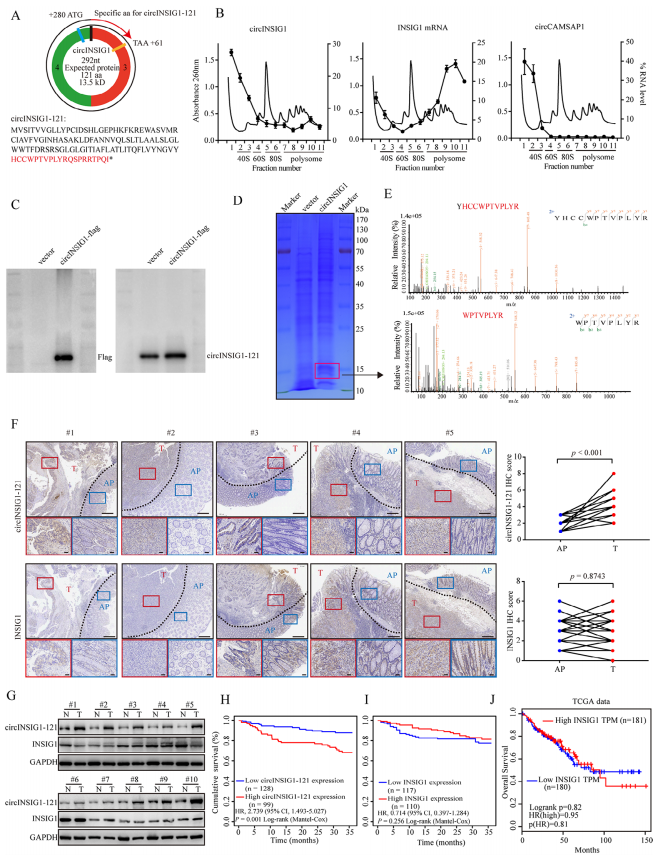
图2 circINSIG1编码121个氨基酸的与CRC的不良预后相关的新蛋白
3、circINSIG1-121促进结直肠癌的增殖和转移
为了阐明circINSIG1在糖尿病中的生物学功能,作者首先利用qRT-PCR分析了circINSIG1在正常人肠上皮细胞系和CRC细胞系中的表达。与此一致,circINSIG1在CRC细胞中被发现上调(图S3A)。由于circINSIG1表达适中,作者选择HCT8和DLD1 CRC细胞系进行细胞实验(图S3A),生成稳定过表达circINSIG1或circINSIG1-121的CRC细胞系(图3A、B和S3B)。球体形成实验证实circINSIG1或circINSIG1-121增强CRC细胞增殖(图3C)。接着在CRC患者源性类器官(PDOs)中过表达circINSIG1或circINSIG1-121,结果也表明circINSIG1或circINSIG1-121促进了PDOs的生长(图3D)。同样,circINSIG1或circINSIG1-121的过表达促进了CRC细胞的迁移和侵袭(图3E和S3C)。作者用两个短发夹RNA敲低了circINSIG1特异性靶向HCT8和DLD1细胞中circINSIG1的后剪接连接的shRNAs(图S3D和S3E)。正如预期的那样,敲低circINSIG1或circINSIG1-121显著抑制CRC细胞的生长、迁移和侵袭(图3F-H和S3F)。此外,构建一个带有起始密码子的突变circINSIG1载体(circINSIG1-ATGmut)。过表达circINSIG1- atgmut不能增强CRC细胞的增殖、迁移和侵袭(图S3G-S3I),提示circINSIG1通过编码circINSIG1-121而不是circINSIG1的环状RNA形式促进CRC细胞的增殖和转移。
接下来,使用原位异种移植肿瘤模型确定circINSIG1-121在CRC进展中的体内功能。结果证实过表达circINSIG1或circINSIG1-121可显著促进肿瘤生长(图3I-J)。对人的分析小鼠肝脏中HPRT mRNA的表达证实了circINSIG1或circINSIG1-121可导致肝转移的肿瘤负荷显著增加(图3 K)。这些结果证明了circINSIG1-121在促进结直肠癌增殖和转移中的关键作用。

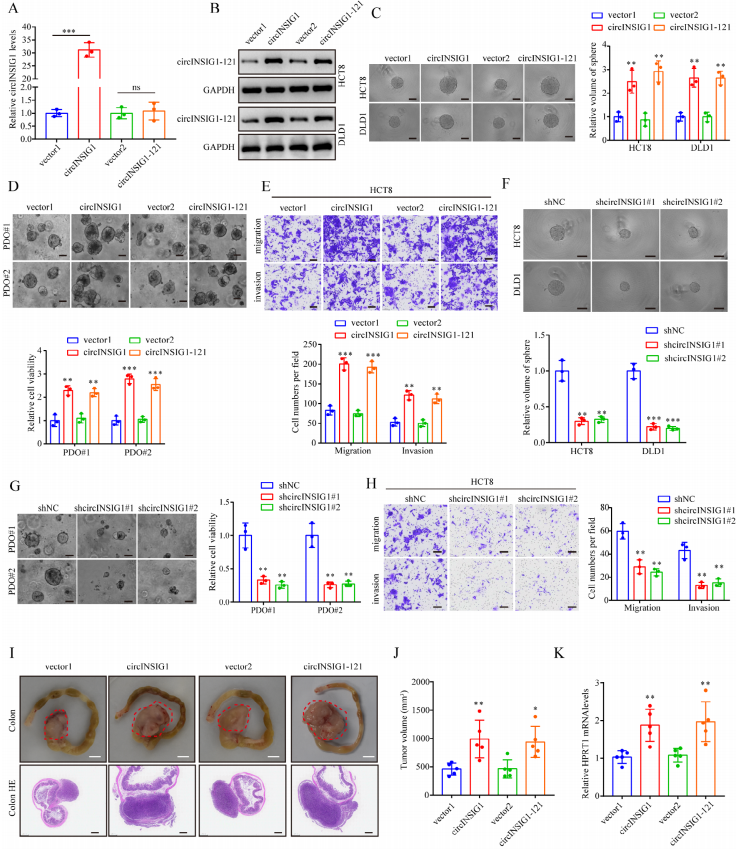
图3 circINSIG1-121促进结直肠癌的增殖和转移
4、circINSIG1-121通过与INSIG1相互作用促进INSIG1不依赖于泛素的降解并诱导胆固醇的生物合成
为了确定circINSIG1-121促进CRC进展的分子机制,作者分析了circINSIG1或circINSIG1-121过表达的HCT8和DLD1细胞中INSIG1的RNA和蛋白水平。有趣的是,INSIG1的RNA水平随着circINSIG1或circINSIG1-121的过表达而升高,而INSIG1的蛋白水平则降低(图4A、B和S4A)。随即通过免疫荧光观察到circINSIG1-121和INSIG1在CRC细胞中的共定位(图4C)。免疫沉淀(IP)实验证实circINSIG1-121与INSIG1相互作用(图4D)。有报道称,INSIG1的磷酸化促进了INSIG1蛋白的降解。因此,作者首先分析了并发现INSIG1的表达磷酸化INSIG1 (p-INSIG1)不因circINSIG1或circINSIG1-121过表达而升高(图S4B)。这些结果表明circINSIG1或circINSIG1-121可能不会通过影响其磷酸化水平来调节INSIG1的降解。由于泛素-蛋白酶体和自噬-溶酶体途径是真核细胞中蛋白质清除的两个主要系统,作者用蛋白酶体抑制剂MG132或自噬-隔离抑制剂3-甲基腺嘌呤(3-MA)处理CRC细胞,以进一步研究circINSIG1-121介导的INSIG1降解的分子机制。我们观察到MG132而不是3-MA提高了INSIG1的蛋白水平,并且在circINSIG1或circINSIG1-121过表达的CRC细胞中,INSIG1的泛素化水平升高,表明circINSIG1-121通过泛素-蛋白酶体途径促进了INSIG1的降解(图4E-F)。
固醇调节元件结合蛋白(SREBPs)在哺乳动物胆固醇和脂质稳态中起着至关重要的作用。INSIG蛋白与与SREBP和SREBP裂解激活蛋白(SCAP)复合物结合,将该复合物锚定在内质网(ER)中,以调节脂肪生成。此外,SREBP1负责脂肪酸合成和能量储存,SREBP2负责胆固醇调节。因此,作者首先分析了全长SREBPs (fSREBPs)和活性核SREBPs (nSREBPs)的蛋白水平。Western blot分析显示,在过表达circINSIG1或circINSIG1-121的CRC细胞中,nSREBP2显著增加,但nSREBP1无显著变化,提示circINSIG1-121可能促进SREBP2激活,从而增强胆固醇的生物合成(图4G)。此外,对SREBP1和SREBP2靶基因进行分析,结果表明SREBP2靶基因表达水平升高,而SREBP1靶基因的表达无明显变化。(图4H和S4C)。与胆固醇生物合成基因的表达增强一致,与对照组相比,circINSIG1-121过表达增加了游离胆固醇含量(图4I)。综上所述,这些数据表明circINSIG1-121促进了INSIG1的泛素依赖性降解,导致内质网释放SREBP2-SCAP复合物,随后诱导胆固醇的生物合成。
为了确认circINSIG1是否以INSIG1依赖的方式重编程胆固醇,进行了体外拯救试验。如图S4D-F所示,过表达circINSIG1可促进CRC细胞增殖、迁移和侵袭,而过表达INSIG1可显著拮抗这一作用。过表达circINSIG1增加了CRC细胞中SREBP2靶基因的表达水平和游离胆固醇的含量,但可以通过过表达INSIG1来下调(图S4G和H)。这些数据表明,circINSIG1通过调节INSIG1的水平,至少在一定程度上重编程胆固醇代谢,促进CRC的恶性进展。
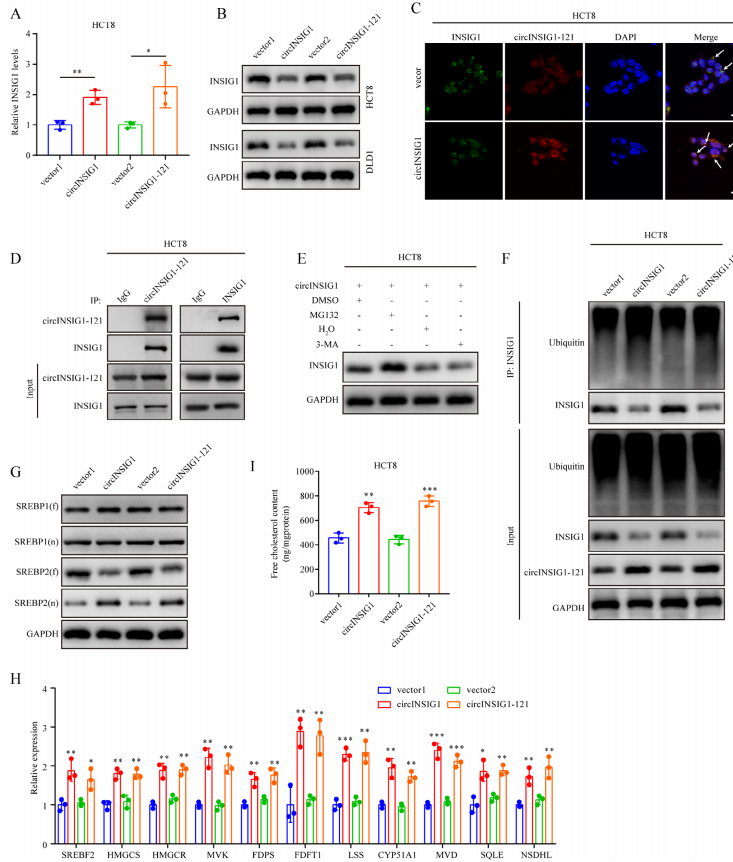

图4 circINSIG1-121促进INSIG1不依赖于泛素的降解并诱导胆固醇的生物合成
5、circINSIG1-121通过招募CUL5-ASB6复合体促进与k48相关的INSIG1泛素化
泛素链是主要的降解信号,负责靶向不同类型的底物,不同种类的泛素链具有不同的生物学功能。为了确定导致INSIG1降解的泛素链类型,我们用不同的HA标记的泛素链载体转染CRC细胞。
研究结果显示,INSIG1主要通过K48连锁泛素化,而不与其他类型的泛素链泛素化(图5A)。在circINSIG1-121过表达的细胞中,k48关联的多泛素化作用增强(图5B)。这些发现表明circINSIG1-121通过k48连锁泛素化介导了INSIG1的降解。然后,作者试图确定INSIG1的哪些赖氨酸残基是导致INSIG1降解的主要泛素化位点,因此,在INSIG1蛋白中鉴定出5个保守赖氨酸(K)点(图S5A),将5个相应赖氨酸残基分别突变为精氨酸(R)的INSIG1突变体分别用于转染的CRC细胞。结果发现,与野生型组相比,转染INSIG1 K156R和K158R组的INSIG1蛋白水平略有下降(图5C-D)。为了进一步证实INSIG1的这两个泛素化位点,构建了具有两个赖氨酸点突变的INSIG1突变体。随后的Western blot和免疫沉淀实验表明,该突变体显著减弱了circINSIG1-121介导的INSIG1降解,且INSIG1的k48连锁泛素化水平显著低于野生型组(图5E-G)。这些结果表明,circINSIG1-121促进了INSIG1在K156和K158残基上的k48连锁泛素化。
由于circINSIG1-121不是E3连接酶,作者推断circINSIG1-121可能作为适配器招募某些E3连接酶使INSIG1泛素化。因此用circINSIG1-121抗体进行免疫沉淀试验,质谱分析鉴定出ASB6的存在(图S5B)。ASB6是一种底物识别适配器,可与Cullin5 (CUL5)相互作用形成CUL5-ASB6复合物,其功能为泛素E3连接酶复合物。进一步的免疫沉淀实验验证了circINSIG1-121和CUL5-ASB6复合物之间的相互作用(图5H-I)。作者发现蛋白酶体抑制剂MG132和Cullin-Ring E3连接酶活性抑制剂MLN4924导致内源性INSIG1蛋白水平明显升高(图5J)。ASB6过表达增强了与k48相关的INSIG1泛素化,降低了INSIG1蛋白的丰度(图5K)。相反,ASB6敲低增加了INSIG1蛋白的丰度,抑制了与k48相关的INSIG1泛素化(图5K)。此外,ASB6的过表达促进胆固醇的生物合成,反之亦然(图5L)。因此,circINSIG1-121招募CUL5-ASB6复合物,促进k48相关的INSIG1在K156和K158的泛素化。


图5 circINSIG1-121促进与k48相关的INSIG1泛素化分析
6、缺氧诱导的EIF4A3促进circINSIG1的表达
作者接着探讨了circINSIG1在结直肠癌细胞中的上调机制。研究表明,RNA结合蛋白(RBP)通过结合环状RNA的侧翼内含子区域参与环状RNA的生物发生。在这里,CircInteractome(探索circRNA及其相互作用蛋白的工具)被用来寻找负责circINSIG1循环化的潜在RBP。在circINSIG1反剪接位点的上游和下游分别鉴定出4个和6个EIF4A3结合位点(图6A和S6A)。EIF4A3是外显子连接复合体的核心组分,在pre-mRNA剪接中发挥重要作用。然后,作者分析了TCGA数据库中EIF4A3的表达,发现EIF4A3在CRC中表达上调(图S6B)。此外,EIF4A3的表达与缺氧标志物HIF-1α的表达呈正相关(图S6C)。Western blot和qRT-PCR检测显示,缺氧条件下EIF4A3蛋白表达上调,circINSIG1表达与EIF4A3表达呈正相关(图6B-C)。为了验证EIF4A3与circINSIG1侧翼序列结合的能力,进行了RNA下拉试验。结果表明,EIF4A3可以与circINSIG1的下游侧翼序列结合,但不是上游序列(图6D和S6D)。然后,我们构建了circINSIG1小基因,并进行了RNA免疫沉淀(RIP)检测。如图6E所示,抗EIF4A3组中6个推测的EIF4A3结合位点的富集程度高于抗IgG组,而转染circINSIG1 miniigene进一步提高了富集程度,表明EIF4A3可以结合到侧翼区域上这些推测的结合位点。我们进一步构建了稳定过表达EIF4A3和一系列具有EIF4A3结合位点突变的circINSIG1小基因的CRC细胞系(图S6E和6F)。值得注意的是,单个推定结合位点的突变对circINSIG1的表达几乎没有影响,而所有六个EIF4A3结合位点的突变大大减少了circINSIG1的形成(图6G)。这些结果表明,这六个位于circINSIG1下游侧翼区域的EIF4A3结合位点是EIF4A3介导的circINSIG1环状化所必需的。此外,我们观察到circINSIG1水平随着EIF4A3过表达而升高,而通过敲低EIF4A3而降低(图6H、I和S6F)。EIF4A3的过表达促进了SREBP2靶基因的激活和胆固醇的生物合成,这可以通过沉默circINSIG1来部分减弱(图6J-L)。总之,这些数据表明缺氧诱导的EIF4A3促进了circINSIG1的生物发生。
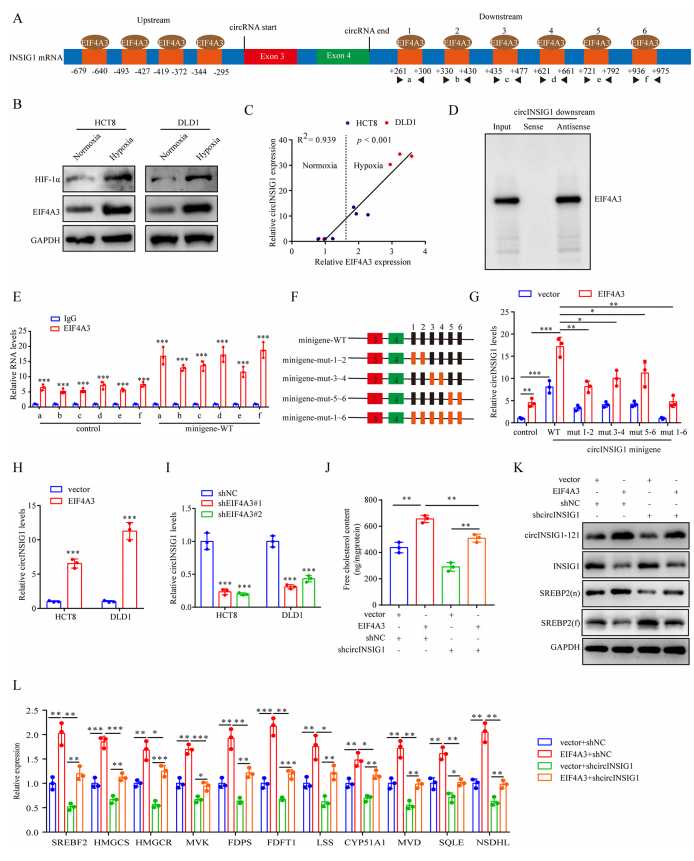
图6缺氧诱导的EIF4A3促进circINSIG1的表达
7、circINSIG1是CRC患者的潜在治疗靶点
为了验证circINSIG1是否可以作为CRC的潜在治疗靶点,我们分析了circINSIG1和circINSIG1-121在6种患者来源的异种移植(PDX)模型中的表达。作者选择circINSIG1上调的PDX3和PDX6靶向circINSIG1检测其治疗效果(图S7A和S7B)。结果显示,shcircINSIG1慢病毒治疗导致肿瘤生长明显低于对照组(图7A-B)。免疫组化分析显示,shcircINSIG1慢病毒治疗组的Ki67阳性细胞百分比明显低于对照组(图7C)。为了进一步验证circINSIG1在体内的作用机制,作者评估了circINSIG1下游靶点的表达并检测了这些肿瘤中的胆固醇含量。结果表明,敲低circINSIG1可降低circINSIG1-121的蛋白水平,抑制INSIG1介导的SREBP1信号传导(图7D-F)。此外,敲低circINSIG1的PDX肿瘤中游离胆固醇含量降低(图7G)。综上所述,这些数据表明circINSIG1是CRC患者的一个新的治疗靶点。
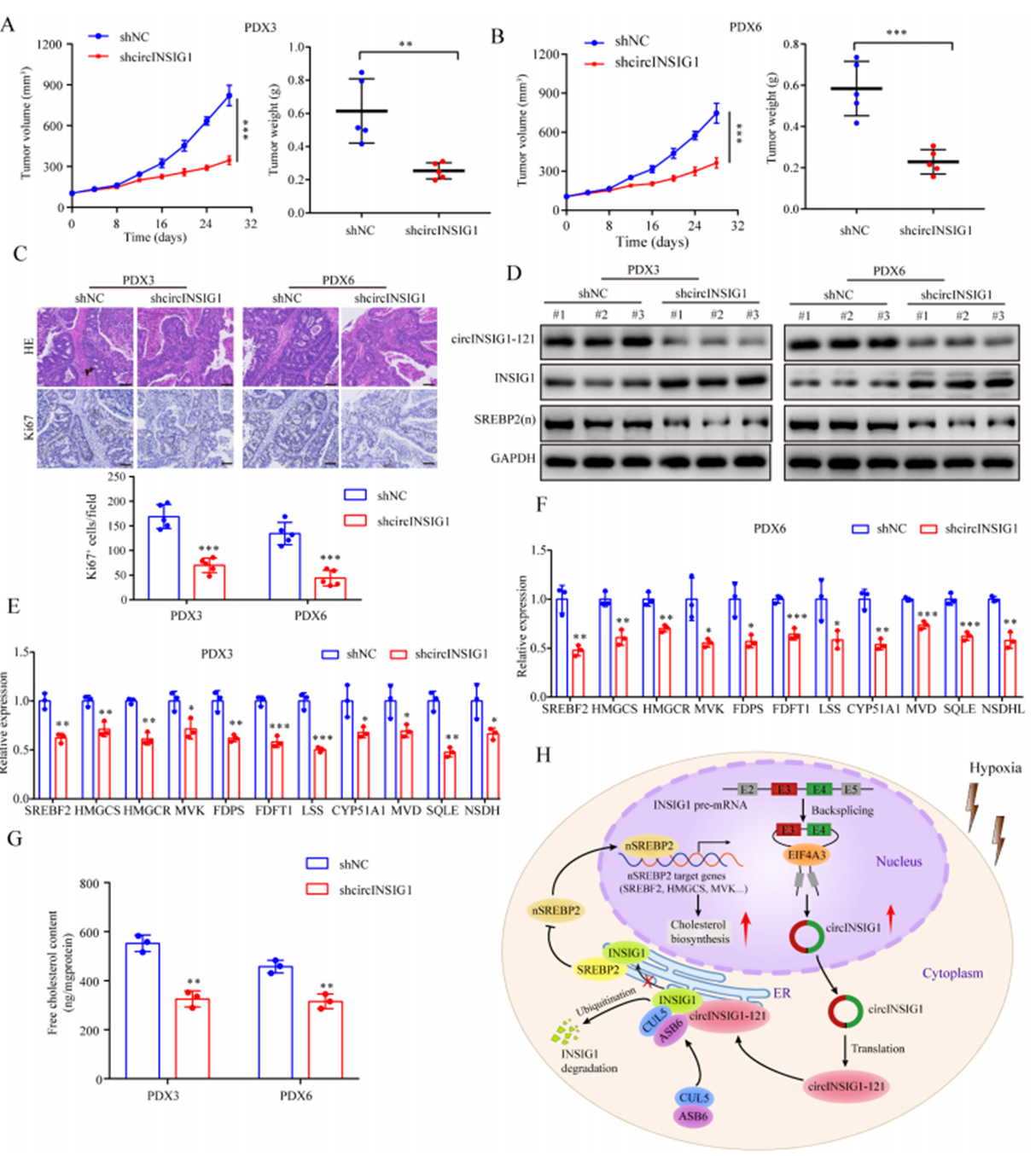
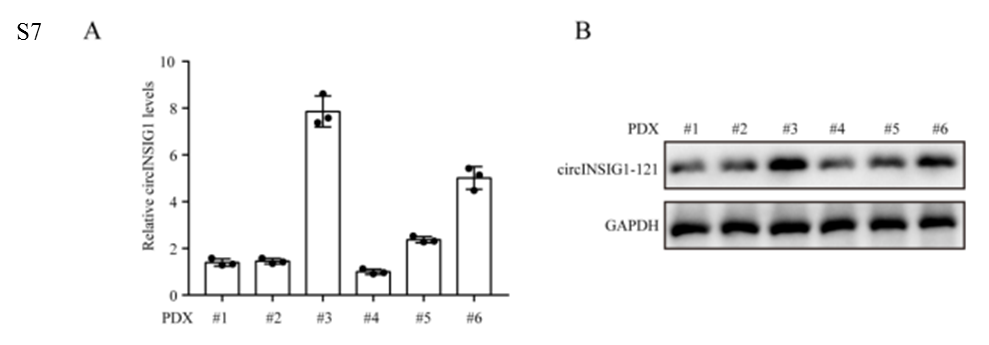
图7 circINSIG1作为CRC患者的潜在治疗靶点分析
实验方法:
高通量RNA测序,RNase R消化实验,qRT-PCR,Western blot,RNA 干扰,过表达,C62H86N12O16测定实验,细胞迁移实验,细胞增殖抑制试验,荧光原位杂交(FISH),原位杂交(ISH),免疫组化(IHC),RNA pull-down,RNA免疫沉淀(RIP)。
参考文献:
Xiong ZL, Liu HS, Zhou C, Yang X, Huang L , Jie HQ, Zeng ZW, Zheng XB, Li WX, Liu ZZ, Kang L, Liang ZX. A novel protein encoded by circINSIG1 reprograms cholesterol metabolism by promoting the ubiquitin-dependent degradation of INSIG1 in colorectal cancer. Molecular Cancer, 2023, April, 22(1):72.
https://doi.org/10.1186/s12943-023-01773-3.
