






抑制糖酵解就可以抑制乳腺癌细胞生长转移!

缺氧依赖型乳腺癌细胞可通过促进糖酵解增强其致瘤性和转移性。N6-甲基腺苷修饰(m6A)是常见的RNA修饰机制。本研究中,作者发现肿瘤缺氧可以转录诱导HIF1α,转录后抑制miR-16-5p的表达,促进YTHDF1的表达,并通过上调m6A修饰的PKM2促进肿瘤细胞糖酵解,最终增加乳腺癌细胞的肿瘤发生和转移潜能。该研究于2022年3月发表在《Cell Death Disease》,IF:9.0。
技术路线
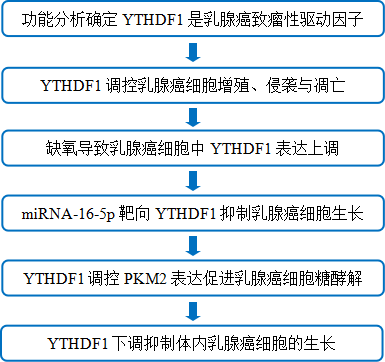
主要研究结果
1. 功能分析确定YTHDF1是乳腺癌致瘤性驱动因子
作者首先使用TCGA和CPTAC数据库分析得到乳腺癌中YTHDF1蛋白和mRNA表达水平显著高于邻近正常组织(图1A和B),114名乳腺癌患者的TCGA数据显示同样结果(图1C)。通过GEPIA2分析发现,YTHDF1高表达与乳腺癌患者的低生存率相关(图1D)。此外,作者收集的22例乳腺癌患者的癌组织及癌旁组织免疫组化结果显示,其中18个样本的癌组织中YTHDF1的表达水平较癌旁组织均有所升高,其余4例差异不明显甚至低于癌旁(图1E)。Western blot结果显示,YTHDF1在乳腺癌组织中表达显著上调(图1F),在MCF10A和多种乳腺癌细胞中YTHDF1的mRNA和蛋白水平呈现同样的上调趋势(图1G和H)。这些结果表明,YTHDF1上调是乳腺癌的一个标志。
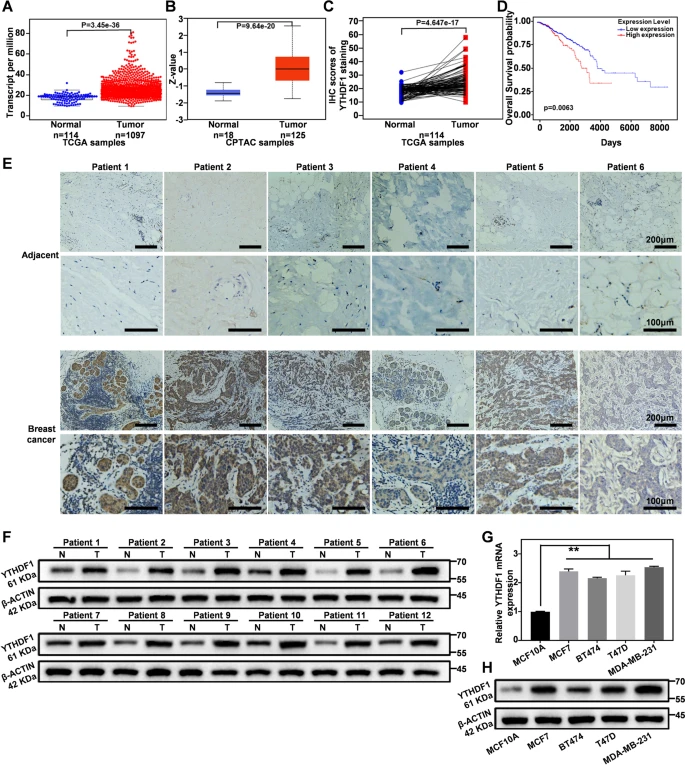
图1:YTHDF1在乳腺癌中上调且与预后不良相关
2. YTHDF1调控乳腺癌细胞增殖、侵袭与凋亡
为进一步阐明 YTHDF1 在乳腺癌中的潜在调节作用,作者通过敲除YTHDF1研究其表型变化。靶向YTHDF1的siRNA转染细胞后,YTHDF1蛋白和mRNA表达水平显着下降(图2A和B)。CCK8检测结果显示,YTHDF1敲低显著抑制MDA-MB-231和MCF7细胞的生长(图2C)。通过EdU染色进一步验证YTHDF1敲低对乳腺癌细胞增殖的抑制作用(图2D)。另外还发现 YTHDF1 敲低抑制乳腺癌细胞集落形成(图 2E)。同时,细胞迁移和侵袭实验表明,YTHDF1敲除后MDA-MB-231和MCF7细胞的迁移和侵袭能力显著下降(图2F)。经Annexin V/PI染色的流式细胞仪检测结果显示,YTHDF1敲低导致MDA-MB-231和MCF7细胞凋亡(图2G)。以上结果表明YTHDF1在维持乳腺癌细胞恶性发展过程中发挥重要作用。

图2:YTHDF1敲低抑制体外细胞增殖与侵袭
3. 缺氧导致乳腺癌细胞中YTHDF1表达上调
缺氧可增强乳腺癌细胞增值与侵袭。为解临床相关条件下YTHDF1在乳腺癌组织中的调控机制,作者进一步研究缺氧对乳腺癌细胞中YTHDF1表达的影响。结果发现,缺氧条件下YTHDF1的mRNA和蛋白表达水平在MDA-MB-231和MCF7细胞中显著上调(图3A和B)。此外,作者将HIF1α和HIF2α siRNA转染MDA-MB-231和MCF7细胞发现,HIF1α基因的缺失导致YTHDF1表达中度降低,而HIF2α的缺失对YTHDF1表达没有明显影响。
缺氧可以通过调节microRNAs来介导特异性标记物的转录后表达。为研究microRNAs可能参与缺氧介导的YTHDF1表达的过程,作者在多个microRNAs数据库搜索筛选了29个可能与YTHDF1相互作用的microRNAs(图3C),并进一步获得参与缺氧介导YTHDF1过表达的10个microRNAs。然后作者发现,在缺氧条件下,miR-16-5p、miR-15a-5p和miR-23b-3p的表达水平显著下降(图3D)。通过转染microRNAs的mimics,结果发现miR-16-5pmimics对YTHDF1 mRNA表达具有最显著的抑制作用(图3E)。为进一步阐明miR-16-5p在缺氧介导的YTHDF1表达中的调控作用,作者构建YTHDF1 3’UTR野生型和突变型的双荧光素酶表达系统(图3F),并与miR-16-5pmimics或miR-16-5p抑制剂共转染。荧光素酶活性结果显示,在YTHDF1 3’UTR野生型中,miR-16-5抑制剂可以逆转miR-16-5p介导的荧光素酶表达活性下调,但不能逆转YTHDF1 3’UTR突变型的荧光素酶活性降低效果(图3G)。以上结果表明,肿瘤缺氧微环境可以通过增强HIF1α的表达以及抑制内源性miR-16-5p的表达来上调YTHDF1。

图3:缺氧诱导YTHDF1在乳腺癌细胞中的表达
4. miRNA-16-5p靶向YTHDF1抑制乳腺癌细胞生长
为确定miR-16-5p介导的YTHDF1抑制对乳腺癌细胞的影响,作者监测miR-16-5pmimics及抑制剂转染乳腺癌细胞后的表型变化。他们首先观察到miR-16-5p可以高靶向消除YTHDF mRNA和蛋白表达。然而,pCDNA3.1-YTHDF1×FLAG质粒与miR-16-5p模拟物共转染可恢复乳腺癌细胞中YTHDF1蛋白表达水平(图4A和B)。同时,CCK8检测结果显示,转染miR-16-5pmimics可显著抑制乳腺癌细胞增殖,而miR-16-5p与miR-16-5p抑制剂或YTHDF1表达质粒联合使用可缓解miR-16-5p介导的抑制作用(图4C和D)。EdU染色结果也支持miR-16-5p对肿瘤的抑制作用和YTHDF1表达质粒的挽救作用(图4E和F)。作者同时也观察到miR-16-5抑制乳腺癌细胞的克隆、迁移和侵袭,而加入miR-16-5p抑制剂或pCDNA3.1-YTHDF1×FLAG质粒则逆转了肿瘤抑制作用(图4G-J)。基于Annexin V/ PI染色的流分析进一步表明,转染miR-16-5p可有效诱导MDA-MB-231和MCF7细胞凋亡,而YTHDF1表达质粒可缓解细胞凋亡(图4K和L)。这些实验结表明,miR-16-5p抑制YTHDF1表达同YTHDF1敲低作用效果一致,均能够抑制乳腺癌细胞的恶性特征和诱导癌细胞凋亡,使miR-16-5p作为靶向调控YTHDF1表达治疗形式的应用成为可能。

图4:miR-16-5p通过下调YTHDF1抑制体外乳腺癌细胞增殖与侵袭
5. YTHDF1调控PKM2表达促进乳腺癌细胞糖酵解
乳腺癌细胞通过糖酵解将大量乳酸终产物释放到肿瘤细胞外微环境中导致其酸化。考虑到缺氧与YTHDF1之间的联系,作者首先研究MDA-MB-231和MCF7细胞中YTHDF1被抑制后,缺氧条件下细胞外微环境的乳酸水平变化。作者发现YTHDF1敲低明显改善缺氧时乳腺癌细胞外微环境酸化,同时显著降低葡萄糖消耗和乳酸产生水平。这说明在乳腺癌细胞中,YTHDF1敲低可以阻断葡萄糖代谢。为进一步阐明YTHDF1在乳腺癌细胞糖酵解调控网络中的作用,作者利用YTHDF1抗体对细胞做了RIP处理,通过qPCR检测糖酵解相关基因在乳腺癌细胞糖酵解调节网络中的表达变化。实验结果显示,PKM2 mRNA被YTHDF1有效富集(图5A)。由于YTHDF1通过识别和结合mRNA的m6A来调控靶基因表达,因此作者在RMBase v2.0数据库中检索了YTHDF1与PKM2 mRNA结合的m6A修饰基序,以确定YTHDF1与PKM2之间的相互作用。利用RAP-CLIP鉴定出PKM2与YTHDF1结合的mRNA m6A修饰基序是ATGGACT(图5B)。关于YTHDF1识别m6A修饰位点后对PKM2的调控机制,作者发现敲低YTHDF1对PKM2 mRNA或蛋白的稳定性没有明显影响。因此,作者推测YTHDF1可能影响PKM2蛋白翻译过程并通过构建靶向PKM2 CDS的双荧光素酶报告系统来验证这一假设。双荧光素酶测定结果显示,与对照组相比(图5C),PKM2在YTHDF1敲低细胞系中的翻译效率降低约40%。这些结果表明,YTHDF1主要通过介导PKM2蛋白翻译过程影响PKM2信号传导。
为进一步研究YTHDF1-PKM2轴在乳腺癌发展过程中的参与作用,作者将YTHDF1敲除后发现,重新表达PKM2可以有效恢复乳腺癌细胞糖酵解活性(图5D-G)。即使YTHDF1过表达,PKM2敲低后仍然抑制糖酵解(图5H-K)。另外,流式细胞仪结果表明,重新表达PKM2可以逆转YTHDF1敲低所诱导的细胞凋亡,而PKM2敲低和YTHDF1过表达联合处理仍导致乳腺癌细胞严重凋亡(图L和M)。这些结果表明,YTHDF1通过调节PKM2的表达影响乳腺癌细胞的糖酵解,从而对乳腺癌细胞的各种生物学活性产生深远影响。

图5:YTHDF1调控糖酵解中PKM2的表达
6. YTHDF1下调抑制乳腺癌细胞体内的生长
为验证YTHDF1在乳腺癌发展中的作用,作者利用皮下乳腺肿瘤的BALB / c小鼠进一步研究YTHDF1敲低对乳腺癌细胞致瘤的影响,并通过向4周龄雌性BALB / c小鼠注射YTHDF1 shRNA处理的4T1细胞构建相关模型。与对照组相比,作者观察到YTHDF1缺失引起肿瘤体积明显减小,细胞凋亡群体增加,并伴随有肿瘤组织中PKM2表达降低(图6A-C),这表明YTHDF1敲低通过抑制PKM2可有效抑制小鼠乳腺肿瘤生长。同时YTHDF1沉默也显著减少小鼠乳腺肿瘤的肺转移(图6J)。此外,作者将YTHDF1过表达转染的4T1细胞转入4周龄雌性BALB / C小鼠中,通过监测肿瘤大小变化来评估YTHDF1过表达对BALB/C小鼠中乳腺癌细胞致癌性的影响。结果发现,与对照组相比,YTHDF1过表达导致皮下乳腺癌组织的大小和PKM2表达水平增加(图6D-F)。同时,YTHDF1过表达也显著增强了乳腺癌细胞的肺转移能力(图6K),具体表现在YTHDF1 过表达处理组转移结节数量增加。这些结果均表明,YTHDF1是介导下游PKM2信号调控乳腺癌发生和转移的正向调节因子。基于以上实验结果,作者利用同样的方法评估了miR-16-5p介导的YTHDF1抑制以及体内抗肿瘤作用。与阴性对照相比,miR-16-5p处理组诱导肿瘤组织中YTHDF1和PKM2的表达显著降低。而且miR-16-5p处理组的肿瘤生长受到明显抑制,给药24天后,平均肿瘤体积大小约为对照组的1/4(图6G-I),其肺转移淋巴结数量远远低于其他两组(图6L)。对提取的肿瘤组织进行Western blot分析,其结果与体外实验结果相似,即YTHDF1缺失抑制了小鼠异种移植肿瘤中PKM2表达。作者通过免疫组化进一步研究患者来源的乳腺肿瘤和邻近正常组织中YTHDF1和PKM2表达水平,结果表明YTHDF1和PKM2在肿瘤组织中均上调,且两者之间呈正相关(图6M)。综上所述, YTHDF1在促进乳腺肿瘤生长和体内转移中起关键作用,而miR-16-5p通过选择性抑制YTHDF1表现出潜在的抗肿瘤作用。
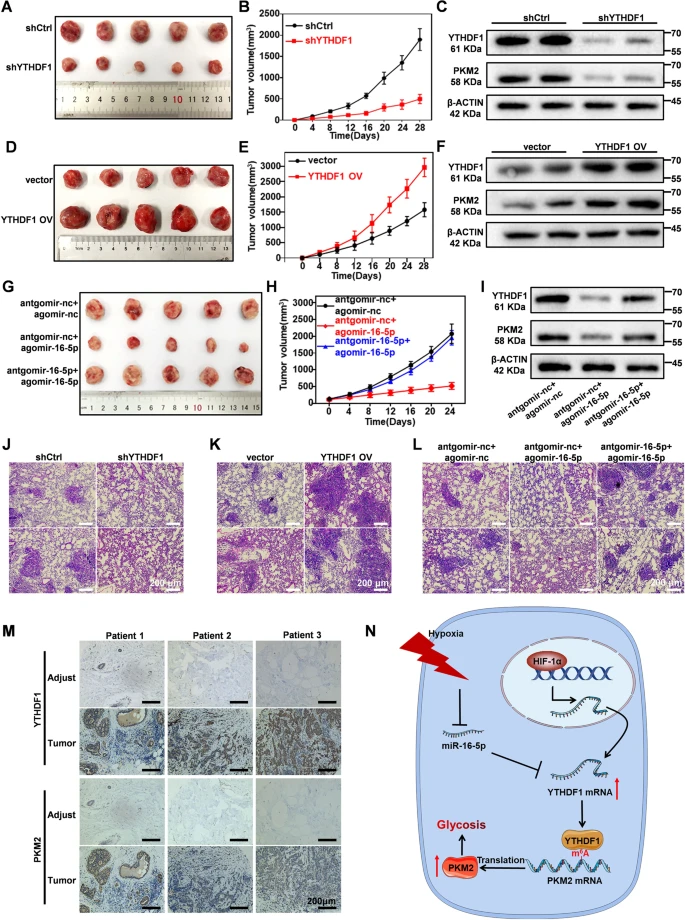
图6:YTHDF1作为小鼠中调控乳腺癌细胞致瘤性和转移性的治疗靶标
结论
综上所述, YTHDF1表达水平上调可以通过调控PKM2蛋白翻译促进糖酵解来增强乳腺癌细胞的增殖和侵袭能力。YTHDF1 介导的 m6A 甲基化修饰可能成为开发更先进的乳腺癌疗法靶标。
实验方法
细胞培养,免疫组化,菌落形成测定,细胞迁移和入侵实验,siRNA 、shRNA或microRNA 转染,CCK8,EdU染色,流式细胞仪检测细胞凋亡,Western blot,RT-PCR,RIP,双荧光素酶报告系统,transwell入侵试验,小鼠动物模型,TCGA、CPTAC数据库分析,葡萄糖和乳酸浓度测定
参考文献
Yao X, Li W, Li L, Li M, Zhao Y, Fang D, Zeng X, Luo Z. YTHDF1 upregulation mediates hypoxia-dependent breast cancer growth and metastasis through regulating PKM2 to affect glycolysis. Cell Death Dis. 2022 Mar 23;13(3):258. doi: 10.1038/s41419-022-04711-1. PMID: 35319018; PMCID: PMC8940925.