






通过分泌组分析鉴定MYH9作为滑膜细胞迁移和侵袭的关键调节因子
类风湿性关节炎(RA)的特征是滑膜增生和广泛的血管生成,称为血管膜形成,这被认为是RA的病理标志。RA患者的纤维原细胞样的滑膜细胞 (RA-FLSs)分泌各种疾病加重因子。这些因素对RA的发病机制至关重要,因为它们在滑膜炎症的持续、免疫细胞的化学吸引、新血管的促进、自身抗原的产生和对关节的长期性损伤中起着重要作用。因此,RA-FLSs分泌的各种分子可以作为有用的生物标志物,代表RA的疾病严重程度,并可能代表治疗靶点。然而,来自RA-FLSs的分泌分子的全局系统概况仍有待澄清。因此,可以将蛋白质组学分析和平行反应监控(PRM)相结合来评估RA-FLS衍生的分泌组,并随后发现与侵袭性血管翳相关的关键分泌蛋白。本文发布于《Annals of the Rheumatoid Arthritis》,IF=27.4。
技术路线
主要研究结果
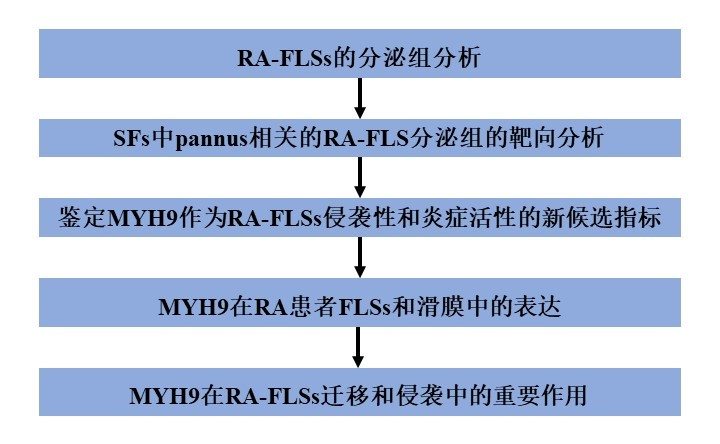
1、RA-FLSs的分泌组分析
为了评估RA-FLSs的促炎分泌组,作者培养了从RA患者滑膜组织中分离的FLSs,这些患者未经治疗(对照组)或接受肿瘤坏死因子(TNF)α+白细胞介素(IL)-1β治疗,模拟RA关节的炎症状态(图1A)。在汇集培养上清后,作者将汇集的样品分成24个部分,并使用液相色谱-串联质谱(LC-MS/MS)分析对单个部分进行蛋白质组学分析(图1A)。使用LC-MS/MS数据集,作者通过使用MS- gf +和UniProt人类参考蛋白质组数据库(图1A)进行数据库搜索,鉴定出843种具有三个以上独特兄弟肽的RA-FLS分泌蛋白,错误发现率<1%。基因本体细胞成分的富集分析表明,843个蛋白中最显著富集的蛋白位于细胞外区域(747个蛋白)和细胞外囊泡(EVs);597个蛋白)(图1B)。此外,843种蛋白中有94.3%与人血浆中检测到的细胞外蛋白或分泌蛋白重叠(图1C)。总的来说,这些数据支持作者通过LC-MS/MS分析获得的RA-FLS分泌组的有效性。接下来,作者使用DAVID软件对基因本体生物过程(Gene Ontology Biological processes, GOBPs)进行富集分析,研究了与RA-FLSs释放的843种蛋白质相关的细胞过程。RA-FLS分泌组主要与代谢(27.4%)、发育过程(19.2%)、信号转导(17.4%)、细胞粘附/迁移(15.7%)、细胞增殖(8.7%)和免疫应答(7.0%)相关(图1D)。值得注意的是,RA-FLS分泌组最强烈地代表了与肠膜形成相关的细胞过程;总体而言,48.5%(409个蛋白)的分泌组与pannus介导的RA病理相关,包括细胞迁移/侵袭(179个蛋白)、细胞外基质组织(177个蛋白)、细胞增殖/凋亡(178个蛋白)、血管生成(46个蛋白)和pannus相关的信号通路(21个蛋白)(图1E)。此外,与信号转导相关的RA-FLS分泌组显著代表了转化生长因子(TGF)β、骨形态发生蛋白(BMP)、Wnt、RhoA和血管内皮生长因子(VEGF)信号转导,以及参与免疫反应的信号转导(核因子κ B、Fc受体和TNF信号转导)(图1F)。因此,RA-FLSs分泌组可以提供一个全面的RA-FLSs分泌因子列表,这些因子参与了pannus的形成,而pannus是RA的病理标志。

图1. RA-FLS分泌组的特征
2、SFs中pannus相关的RA-FLS分泌组的靶向分析
为了进一步表征RA-FLSs的分泌组谱,作者使用PRM方法进行了靶向蛋白定量,并试图确定临床相关性。简单地说,作者从843个蛋白中选择了493个参与pannus相关的细胞过程(图2A)。其中,作者根据前面描述的标准,选择了151个含有2或3个定量肽的蛋白质,并获得了高质量的MS/MS谱,然后作者构建了包含151个蛋白质的436个定量肽的MS/MS谱的谱库(见图1A, pannus相关分泌组)。为了测试151个蛋白的生物学相关性,作者对SFs(关节中的代表性生物液)中的151个pannus相关分泌蛋白进行了PRM分析,SFs来自RA (n=117)和骨关节炎(n=45,非RA对照)患者,通过关节穿刺获得。在使用IgY14耗尽柱去除14个丰富的蛋白后,使用PRM分析对单个SF样品中的151个蛋白进行定量(图2B)。在至少一个SF样品中鉴定出对应121个蛋白的277个定量肽的丰度。接下来,作者研究了121种pannus相关分泌蛋白中哪一种与RA的病理严重程度相关,包括滑膜增生和血管生成,在SF取样时通过肌肉骨骼US同时评估;由于US能以无创的方式密切反映关节病变的严重程度,因此被广泛用于RA的研究。因此,作者采用US比较了OA(病理对照)、轻度RA和重度RA三组患者中PRM肽的丰度。根据以下严重程度标准(图2C),作者发现与OA和轻度RA组相比,DEPs在重度RA组中表达增加:(1) 9个DEPs(膜联蛋白A1 (ANXA1), rho GDP解解抑制剂α (ARHGDIA),环化酶相关肌动蛋白细胞骨架调节蛋白1 (CAP1),胶原V型α 2链(COL5A2),纤维连接蛋白1 (FN1),葡萄糖-磷酸异构体酶(GPI), moesin (MSN), MYH9和S100钙结合蛋白A6 (S100A6)),灰色US (GSUS)等级为0和1,GSUS等级为2和3(轻度和重度滑膜肥厚);(2)10个DEPs (ANXA1, CAP1, COL5A2, FN1, GPI, MSN, MYH9, MYH9,帕金森病相关脱糖酶7 (PARK7), S100A6和超氧化物歧化酶2 (SOD2))在功率多普勒US (PDUS)分级为0与PDUS分级为1-3之间的差异。此外,当活动性滑膜炎被定义为GSUS分级为2或3或PDUS分级为1-3时,与非活动性滑膜炎患者相比,活动性滑膜炎RA样本中鉴定出9个DEPs (ANXA1、ARHGDIA、CAP1、COL5A2、FN1、GPI、MSN、MYH9和S100A6)(图2C)。8种典型dep的差异表达反映滑膜肥大、血管扩张和活动性滑膜炎,如图2D所示。作者还试图确定121种蛋白与RA炎症活性的相关性。随后,通过SF WBC计数≥5000×103 cells/mL与SF WBC <5000×103 cells/mL的比较,作者鉴定了10个DEPs (APOA1、arhdia、CAP1、cofilin 1 (CFL1)、GPI、谷胱甘肽s -转移酶P1 (GSTP1)、MSN、MYH9、硫氧还蛋白依赖性过氧化物还原酶(PRDX3)和S100A9),并通过血液CRP水平≥0.5 mg/dL与<0.5 mg/dL的比较鉴定了5个DEPs (GPI、GSTP1、MSN、MYH9和S100A9)(图2E、F)。作者最终选择了16例dep,这些dep与上述四项RA严重程度(基于US)和炎症活性(基于SF WBC计数和血液CRP水平)中的至少一项密切相关。在16种DEPs中,APOA1、FN1、GPI和S100A9作为自身抗原、炎症介质、增殖诱导剂或细胞凋亡抑制剂与RA发病有关。此外,已知某些DEPs是由RA-FLSs产生的(GPI和S100A9),其他DEPs参与RA-FLSs的增殖(MSN)和侵袭(ANXA1和CFL1),并参与炎症细胞因子的产生(CAP1),这表明作者的PRM分析结果与之前的研究结果一致。总的来说,PRM分析鉴定出16个关键dep,表明RA的“侵袭性pannus”和“炎症活性”,随后被称为“滑膜细胞分泌组特征”(SSS)。
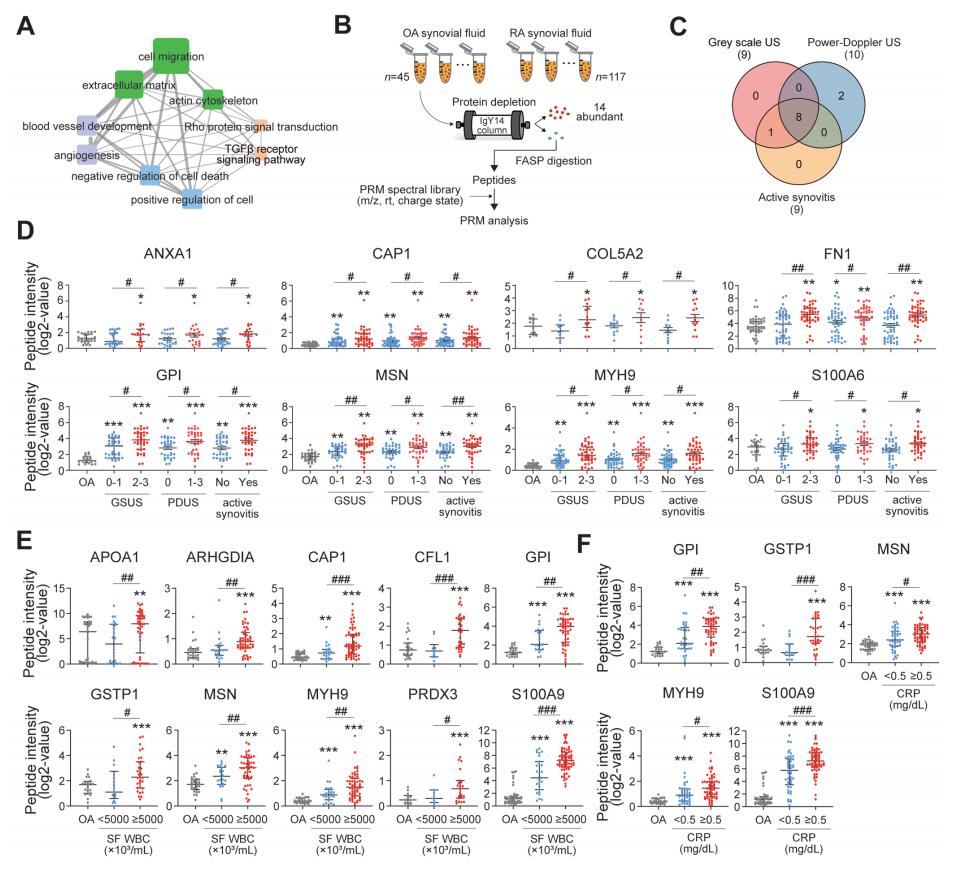
图2. pannus相关RA-FLS分泌组蛋白的差异表达
3、鉴定MYH9作为RA-FLSs侵袭性和炎症活性的新候选指标
在SSS中,GPI、MYH9和MSN三个蛋白满足选择SSS的所有标准(图3A)。虽然GPI和MSN与RA的发病机制有关,但MYH9在RA中的作用很少被研究。因此,作者决定进一步研究这个新的靶点。作者首先使用ELISA比较了RA和OA sf中的MYH9水平,发现RA sf中的MYH9水平明显高于OA sf(图3B)。此外,RA-FLSs产生的主要趋化因子和细胞因子CCL2、IL-6、IL-8、TGFβ和IL-1β的水平,以及RA-FLSs的中枢激活因子TNFα的水平,与RA SFs中MYH9的水平相关(图3C)。这些发现与作者的PRM分析结果一致,显示MYH9水平的升高取决于滑膜炎和全身炎症的严重程度(图2D,F)。类风湿关节炎是一种异质性疾病,涉及多种免疫细胞和细胞过程,其主要病理,尽管存在泛膜形成的共性,但可能因疾病分期和表型而异。作者质疑MYH9表达是否特别反映在优先表现为“侵袭性血管翳”病理的亚组中。因此,作者首先获得了先前报道的使用152例RA患者滑膜组织生成的大量RNA-seq数据集,并使用正交非负矩阵分解聚类(图3D)确定了RA的三个主要亚型(Sub1, Sub2和Sub3)。为了进一步表征每种亚型的特征,作者试图通过使用ConsensusPathDB进行基因集富集分析来定义每种亚型中主要上调基因所代表的主要细胞通路。Sub1主要与细胞增殖相关的细胞通路相关(如图3E中的细胞周期和G2/M检查点),Sub2与细胞代谢相关的通路相关(如图3E中的丙酮酸、氨基酸和脂质代谢),Sub3与细胞迁移和侵袭相关的通路相关(如图3E中的胶原形成、细胞外基质-受体相互作用和TGFβ/BMP/Hippo信号传导),表明Sub3是侵袭性肠腺所特有的。值得注意的是,MYH9在Sub3中表达水平最高(图3F),这与作者的PRM分析结果一致,表明MYH9与促迁移和促侵袭病理的关系比与其他细胞过程(细胞增殖和代谢)的关系更大。作者进一步分析了从大量RNAseq数据中鉴定的三种亚型滑膜组织中富集的细胞类型。然后,作者通过使用CIBERSORTx对相应的大量RNA-seq数据进行细胞型反褶积,估计了三个亚组的每个滑膜组织中五种细胞类型的比例。作者发现,相对于其他四种免疫细胞类型的比例,Sub3含有高比例的成纤维细胞,与RA-FLSs相对应(图3G)。这些数据,再加上Sub3中MYH9表达的增加(图3F),表明MYH9与RA滑膜中成纤维细胞显性病理密切相关,这与图2D的US分析结果一致。
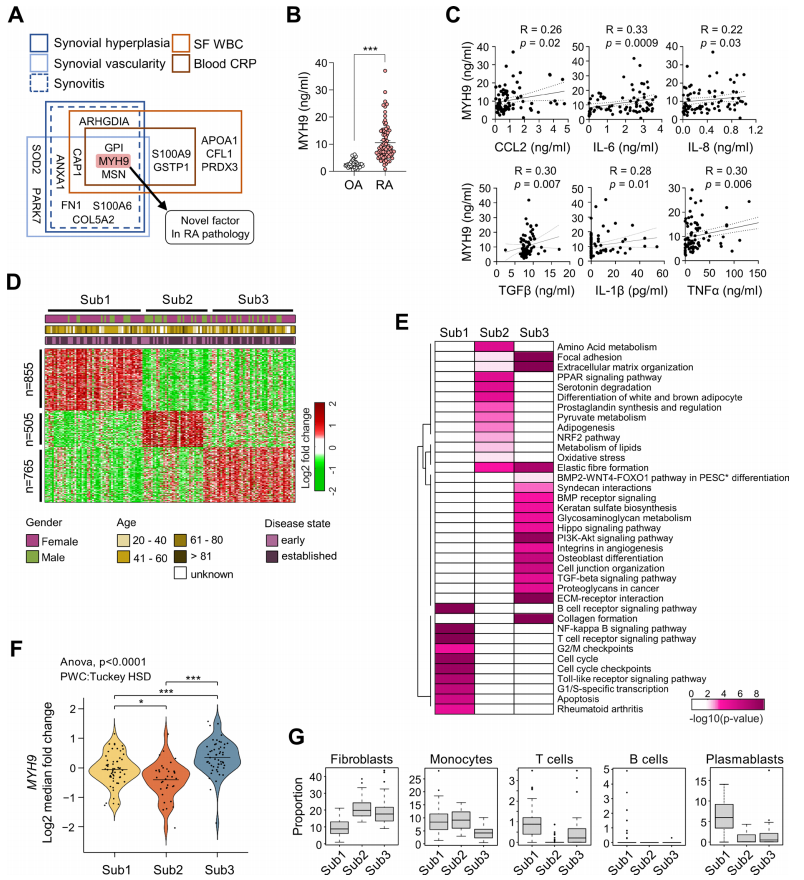
图3. MYH9作为SSS的重要成员,代表RA-FLSs的侵袭性
4、MYH9在RA患者FLSs和滑膜中的表达
基于PRM和细胞型反褶积分析,作者验证了MYH9是否真的在RA-FLSs中表达,是否参与了FLS的迁移和侵袭。正如预期的那样,基于qPCR分析,MYH9 mRNA在RA-FLSs中表达,并且在促炎细胞因子(包括IL-1β和TGFβ)的刺激下表达增加(图4A)。经ELISA检测,IL-1β和TNFα刺激的RA-FLSs分泌MYH9蛋白相似,但TGFβ不分泌MYH9蛋白(图4B)。作者之前证明,RA-FLSs严重暴露于内质网(ER)刺激,如缺氧和促炎刺激。值得注意的是,用内质网应激源tunicamycin治疗RA-FLSs,可显著增加MYH9分泌,而不影响MYH9 mRNA水平(图4A,B);另一种内质网应激源thapsigargin未能诱导MYH9分泌(图4B)。有趣的是,TGFβ和IL-6处理并没有激发RA-FLSs(细胞外)MYH9的分泌(图4B),但通过western blot分析,它确实大幅上调了RA-FLSs(细胞内)MYH9的表达(图4C)。这些数据表明,促炎细胞因子IL-1β和tnf - α以及tunicamin可以诱导RA-FLSs分泌MYH9,并且似乎与其在细胞中的表达水平并不一定相关。RA滑膜细胞不仅由FLSs组成,还包括巨噬细胞样滑膜细胞(MLSs)。从血液中募集的mls和单核细胞是风湿性关节炎关节慢性炎症长期存在所必需的。作者发现,LPS处理后,人血源性单核细胞MYH9 mRNA表达和MYH9蛋白分泌均显著增加(图4D,E)。TNFα也适度增加单核细胞MYH9 mRNA的表达,但不能像LPS那样显著提高MYH9的分泌(图4D,E)。值得注意的是,培养单核细胞的MYH9分泌量明显高于RA-FLSs(见图4B和图E)。此外,通过western blot分析,LPS、TNFα和IL-1β刺激外周单核细胞显著增加(细胞内)MYH9表达(图4F)。这些数据表明,TLR4结扎和促炎细胞因子TNFα和IL-1β可以增加人单核细胞MYH9的分泌和/或表达。免疫组织化学显示,MYH9在RA滑膜中的表达高于OA滑膜,尤其是在粘膜层和下层白细胞中(图4G)。免疫荧光染色同样显示myh9表达细胞与抗cd55 +和antid68 +细胞明显共定位,证实了FLSs和MLSs是RA滑膜中主要表达myh9的细胞(图4H)。总的来说,MYH9在RA滑膜的FLSs和MLSs中表达,并且可以在促炎细胞因子、toll样受体激动剂(LPS)或内质网应激的刺激下从这些细胞分泌(或排泄)。

图4. MYH9在RA- FLSs和RA滑膜中的表达
5、MYH9在RA-FLSs迁移和侵袭中的重要作用
MYH9编码非肌球蛋白IIA重链。由MYH9和MLC组成的非肌球蛋白IIA是细胞粘附和迁移不可缺少的因子。由于FLSs的迁移和侵袭增强是RA的关键病理之一,作者研究了MYH9在这些细胞过程中的作用。作者发现,在TNFα和TGFβ处理的RA-FLSs中,MLC磷酸化(非肌肉肌球蛋白可以转化为活性形式)显著增加,这表明促炎细胞因子刺激增强了RA-FLSs的myh9依赖性活性(图5A)。众所周知,激活的非肌球蛋白IIA通过其头部结构域与f -肌动蛋白结合。在细胞扩散实验(图5B)中,作者发现,在将悬浮的RA-FLSs附着在涂有纤维连接蛋白的玻璃上20min后,MYH9弥散分布在细胞质中,而F-actin主要分布在细胞边缘,其中肌动蛋白细胞骨架经历了广泛的重塑,这是通过F-actin和MYH9的双重免疫染色确定的。随着时间的推移,在RA-FLSs在纤维连接蛋白涂层玻璃上孵育60min和120min后,MYH9主要在细胞边缘被检测到,并与f -肌动蛋白共定位。此外,在RA-FLSs稳定附着一夜后,大多数MYH9在细胞质中弥散重新分布,但在TGFβ刺激下,一些MYH9分子在细胞边缘被发现,并与肌动蛋白细胞骨架中的F-actin共定位(图5C)。总之,这些结果支持了先前的报道,即MYH9活性可以由TGFβ诱导,并与肿瘤细胞中的f -肌动蛋白结合。为了确定MYH9对RA-FLSs迁移和侵袭的影响,作者使用siRNA敲低MYH9(图6A)。敲除MYH9后,RA-FLS迁移(图6B)、实时伤口迁移(图6C)、侵袭(图6D)均显著降低。此外,含有板足的RA-FLSs的数量通过MYH9敲低相应减少(图6E),板足是众所周知的亚细胞结构,代表迁移细胞的肌动蛋白重组。与此同时,MYH9缺陷显著损害了paxillin磷酸化(图6F),这是局灶黏附的重要步骤,也是细胞迁移动力学不可或缺的。综上所述,这些观察结果以及早期的报告表明,TGFβ刺激可以增加MYH9的活性和表达,从而诱导paxillin的磷酸化以及MYH9与F-actin的结合,最终促进RA-FLSs的粘附、迁移和侵袭。为了进一步研究MYH9在体内RA-FLS侵袭性中的作用,作者建立了SCID小鼠异种移植模型,这是一种人源化滑膜炎模型,在SCID小鼠的左侧植入含有RA-FLS的人软骨,在同一小鼠的右侧植入不含RA-FLS的人软骨。植入MYH9缺陷RA-FLSs的软骨在同侧(左侧)和对侧(右侧)的破坏和降解水平均显著降低(图6G),表明MYH9敲低抑制了RA-FLSs的局部侵袭,并可能延缓RA-FLSs从受影响关节向未受影响软骨的远处迁移。体外和体内使用siRNA的研究结果强烈表明,MYH9是调节FLS迁移和侵袭的一个极好的靶点。因此,作者测试了blebbistatin(一种特异性MYH9抑制剂,可抑制非肌球蛋白II的活性)是否能在体内逆转软骨破坏,并在体外抑制RA-FLSs的亲迁移和亲侵袭表型。blebbistatin处理后,RA-FLSs的迁移、侵袭、板壁形成和实时创面迁移均被显著抑制,体外不影响细胞活力(图7A-D),这与MYH9 siRNA敲低实验结果一致。此外,在SCID小鼠体内人源性滑膜炎模型中,腹腔注射blebbistatin (10 mg/kg,每周两次)可显著降低RA-FLSs介导的同侧和对侧软骨降解(图7E)。这些结果表明,blebbistatin抑制了RA-FLSs在同侧的局部侵袭,并阻止了它们向未植入RA-FLSs的对侧迁移。为了在更复杂的关节炎模型中进一步证实这些发现,除了FLSs外,还有多种免疫细胞参与疾病进展,作者引入了两种不同的慢性炎性关节炎小鼠模型,包括甲基化牛血清白蛋白(BSA)/ il -1β诱导的关节炎和胶原诱导的关节炎。结果显示,腹腔注射blebbistatin(每天10 mg/kg)可显著减少BSA/ il -1β诱导的关节炎小鼠的骨和软骨破坏,尽管其对受影响关节的炎症细胞浸润的影响不大(图7F);大多数小鼠耐受blebbistatin甚至重复治疗,并且没有明显的不良反应或毒性,包括血小板减少症(数据未显示)。用blebbistatin (10 mg/kg,每周2次)治疗的小鼠胶原诱导关节炎的严重程度也明显低于单独使用vehicle的小鼠。这些数据表明,blebbistatin治疗可有效改善多种免疫/炎症细胞对关节的破坏,并延缓主要由RA-FLSs介导的关节破坏和疾病传播。综上所述,这些数据证实了MYH9对于RA-FLSs的迁移和侵袭至关重要,并证明了blebbistatin抑制MYH9在体外和体内成功地抑制了RA-FLSs介导的局部侵袭和软骨降解。
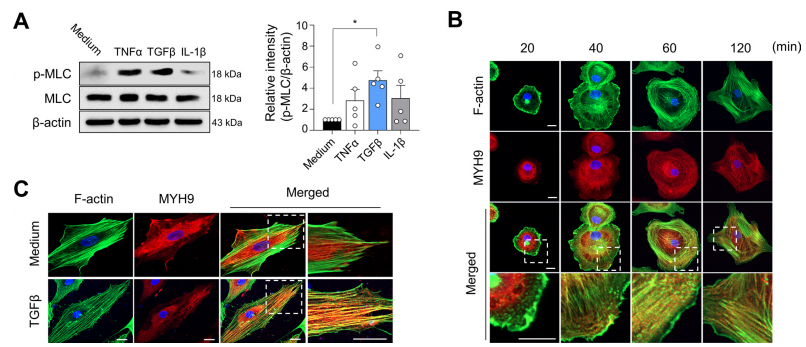
图5. TGFβ对MYH9的激活及其与RA-FLSs中F-actin的共定位
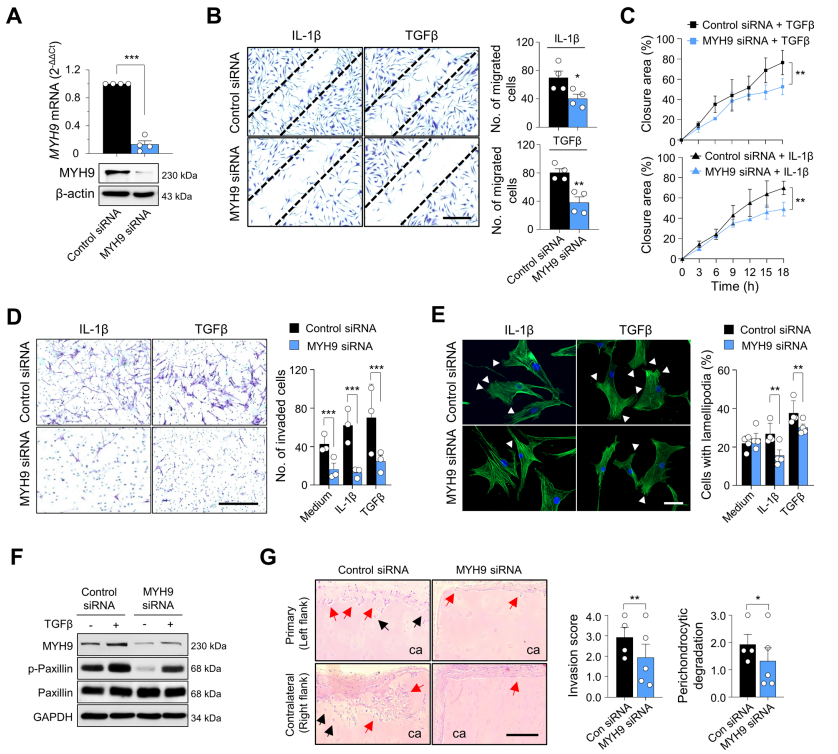
图6. MYH9对RA-FLSs迁移和侵袭的影响
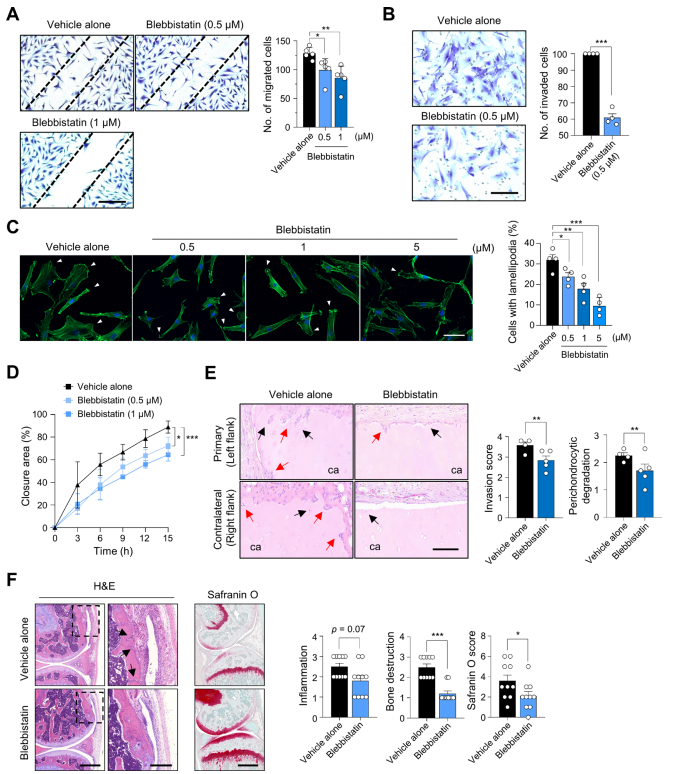
图7. 非肌球蛋白II特异性抑制剂blebbistatin减少RA-FLSs的迁移和侵袭
结论
总之,作者的研究为RA-FLSs衍生的分泌组提供了一个全面的资源,可以应用于各种研究,以帮助发现由RA-FLSs分泌的蛋白质介导的病理过程的新调节因子。此外,对RA-FLSs衍生的分泌组进行PRM分析,发现包含SSS的16种分泌蛋白代表了侵袭性输卵管的病理。类似的PRM分析应该应用于识别RA发病机制中其他过程的关键分泌蛋白,包括RA-FLSs的代谢改变、异常增殖和去分化,因为它成功地分类了侵袭性泛膜相关的SSS。最后,作者提出MYH9是一个有希望延缓RA-FLSs异常迁移和侵袭的靶点,也提出了SSS的潜在治疗候选物,可以设计详细的功能实验。
实验方法
样本收集(滑膜液)、评估滑膜炎严重程度、细胞活力测定、免疫组化、ELISA、q-PCR、WB、分泌组的蛋白质组学分析、基于RNA-seq数据的RA亚型鉴定、RA-FLS分泌组的PRM分析、滑膜组织免疫荧光染色、Cell-spreading化验、细胞侵袭迁移实验、SCID小鼠人源化滑膜炎体内模型
参考文献
Lee S, Choi E, Chae S, Koh JH, Choi Y, Kim JG, Yoo SA, Hwang D, Kim WU. Identification of MYH9 as a key regulator for synoviocyte migration and invasion through secretome profiling. Ann Rheum Dis. 2023 Aug;82(8):1035-1048. doi: 10.1136/ard-2022-223625. Epub 2023 May 15. PMID: 37188496; PMCID: PMC10359537.