






m7G tRNA修饰通过激活SHAF4依赖的线粒体氧化磷酸化促进胃癌进展
m7G tRNA修饰与肿瘤的发生和发展密切相关。然而,m7G tRNA修饰在胃癌(GC)中的确切功能和分子机制尚不清楚。在本研究中,我们评估了METTL1和WDR4在GC中的表达和功能,并阐明了METTL1/WDR4介导的m7G tRNA修饰在促进GC进展中的机制。胃癌组织中m7G甲基转移酶复合蛋白METTL1和WDR4的上调与患者预后不良显著相关。在功能上,METTL1和WDR4在体外和体内促进GC进展。在机制上,METTL1敲减降低了m7G修饰tRNA的表达,并减弱了富含氧化磷酸化途径相关癌基因的翻译。此外,METTL1通过促进SDHAF4的翻译,增强了ETC II的活性,从而加速了GC的代谢和进展。强制表达SDHAF4和化学调节ETC II可以逆转METTL1对小鼠GC的影响。总的来说,我们的发现揭示了METTL1/WDR4介导的m7G tRNA修饰在GC进展中的致癌作用和分子机制,表明METTL1/WDR4及其下游信号轴可能是GC治疗的潜在靶点。本文于2025年3月发表于“Cancer Letters”(IF=9.1)上。
技术路线:

结果:
1)METTL1和WDR4在GC中升高并与不良预后相关
为了探讨m7G tRNA修饰在癌症中的潜在作用,我们使用TCGA数据库分析了METTL1和WDR4在不同癌症中的表达。在多种肿瘤中检测到METTL1和WDR4水平的升高,特别是在胃癌样本(TCGA-STAD)中(图1A)。这些发现在我们临床队列中得到验证,与周围正常组织相比,胃癌组织中METTL1、WDR4和m7G的表达升高(图1B和C)。此外,免疫组化染色也证实了与周围组织相比,肿瘤组织中METTL1和WDR4的表达增加(图1D)。同时,数据库分析和Co-IP实验揭示了胃癌中METTL1和WDR4表达水平之间的正相关关系,以及它们之间的直接结合,表明这两种蛋白质之间可能存在功能性的相互作用(图1E)。为了探索METTL1和WDR4的临床相关性,我们分析了Kaplan–Meier Plotter数据库中的数据,发现METTL1和WDR4表达水平的升高与总生存期(OS)缩短相关(图1F)。此外,我们使用包含94个原发性胃癌组织的组织微阵列进行了免疫组化染色,以评估METTL1/WDR4表达;根据IHC染色评分,将这些样本分为低表达和高表达组(图1G)。Kaplan–Meier生存分析显示,在我们队列中,METTL1或WDR4高表达组的OS比低表达组差(图1H)。这些结果突显了METTL1和WDR4在胃癌进展中的临床重要性。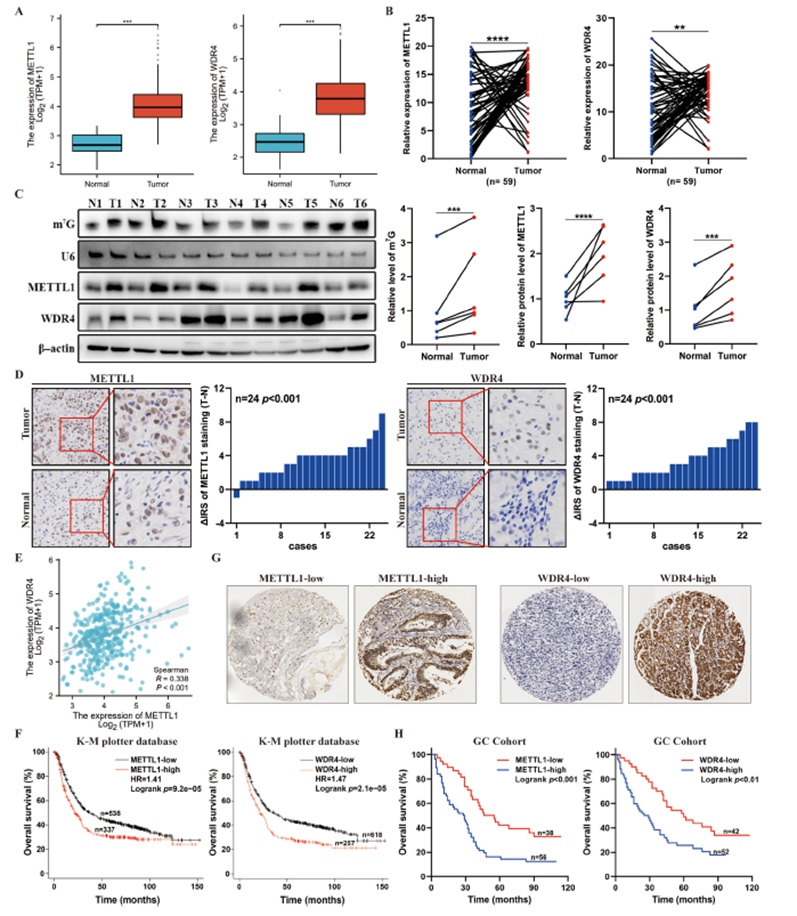
2)METTL1敲除抑制体外和体内GC进展
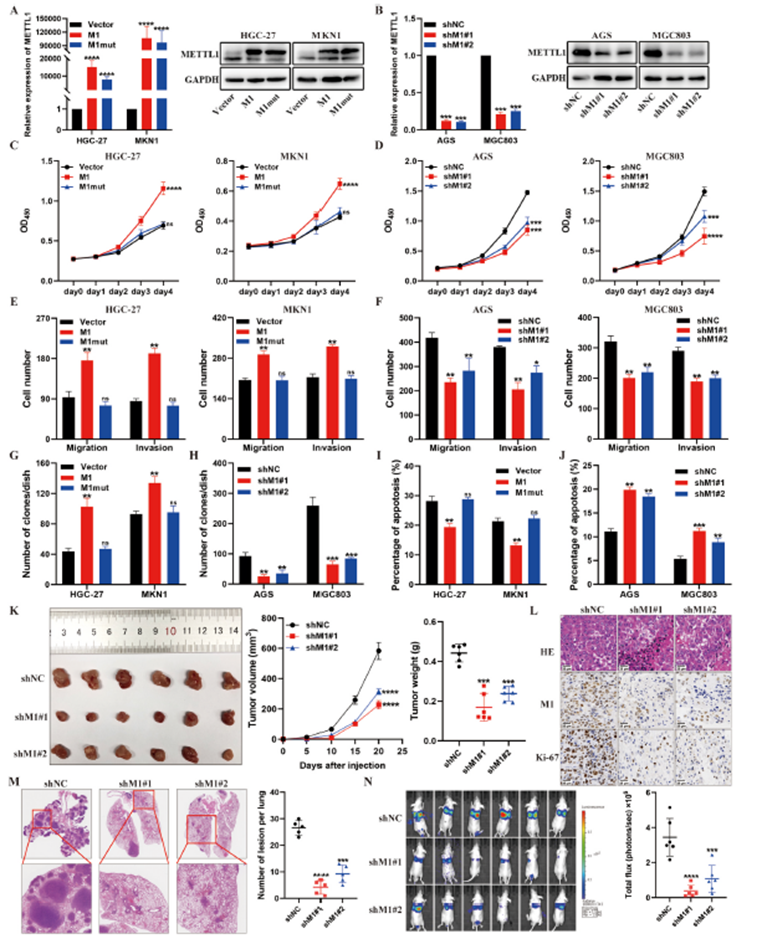
为了研究METTL1在胃癌进展中的生物学功能,我们将METTL1及其催化失活突变体(缺乏m7G tRNA甲基转移酶活性)引入METTL1表达较低的胃癌细胞(HGC-27和MKN1),并通过qRT-PCR和western blotting评估了其效率(图2A和B)。我们发现,野生型METTL1的过表达增强了胃癌细胞的增殖、迁移、侵袭、克隆形成和抗凋亡能力,而其催化失活突变体则没有这种效果(图2C,2E,2G,2I)。作为补充,我们使用两个短发夹RNA(shRNAs)来下调METTL1表达较高的胃癌细胞(AGS和MGC803)中的METTL1。结果表明,METTL1的敲减抑制了胃癌细胞的增殖、迁移、侵袭、克隆形成和抗凋亡能力(图2D,2F,2H,2J)。随后,我们使用异种移植小鼠模型确认了METTL1在胃癌中的体内作用。来自METTL1敲减MKN45细胞的肿瘤生长较慢,其体积和重量显著低于阴性对照组细胞(图2K)。此外,免疫组化染色显示METTL1敲减组的肿瘤组织中Ki-67水平降低,表明METTL1敲减减少了体内胃癌的增殖活性(图2L)。为了研究METTL1对血液转移性胃癌的影响,我们通过将MKN45-shNC或MKN45-shM1细胞注射到裸鼠的尾静脉中,建立了小鼠肺转移模型。METTL1敲减组在注射后40天的转移结节数量和大小显著低于阴性对照组(图2M,2N)。总的来说,这些发现表明METTL1在体外和体内促进了胃癌的进展。
3)METTL1/WDR4调控GC中tRNA m7G修饰、tRNA表达和全局mRNA翻译
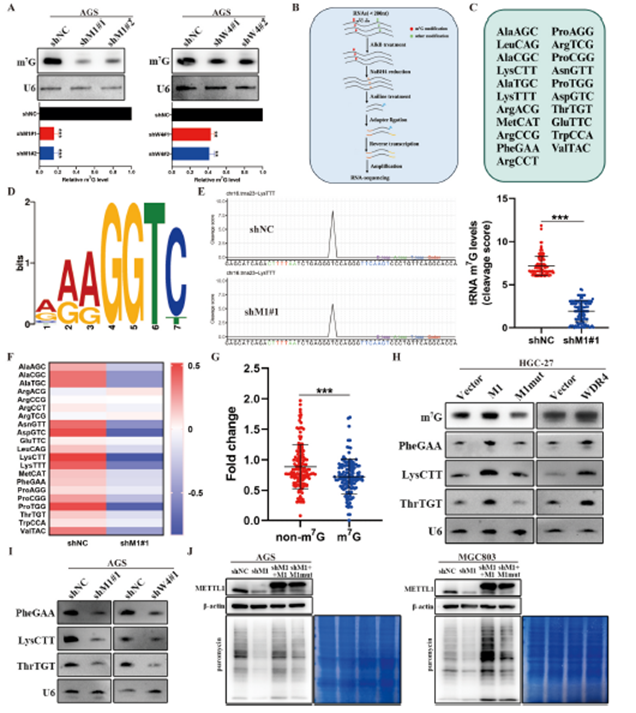
为了探究METTL1/WDR4在胃癌中致癌作用的分子机制,我们进行了Northwestern印迹分析,以评估METTL1和WDR4敲减的AGS细胞中tRNA的m7G信号。结果表明,METTL1或WDR4的敲减降低了tRNA的m7G修饰(图3A)。随后,我们分析了METTL1敲减和阴性对照AGS细胞中全局tRNA m7G修饰的情况,并识别出在可变环中具有"VRRGGTY"基序序列的21种m7G修饰的tRNA(图3B–D)。我们发现METTL1敲减导致这些识别出的tRNA的m7G信号和表达显著降低,表现为较低的切割评分(图3E)。此外,METTL1敲减显著减少了大多数m7G修饰的tRNA的表达,而未修饰的tRNA不受影响(图3F和G)。m7G Northwestern和Northern印迹分析也确认了在METTL1敲减的胃癌细胞中,几种代表性的m7G修饰的tRNA表达下调(图3I)。相反,野生型METTL1的过表达(而非其催化失活突变体)上调了全局tRNA m7G修饰和几种代表性的m7G修饰的tRNA的表达水平(图3H),验证了METTL1通过其m7G甲基转移酶活性调节tRNA修饰和表达。WDR4在胃癌细胞中的敲减或过表达产生了类似的效果(图3H和I)。总的来说,这些结果证明了METTL1和WDR4在tRNA m7G修饰和表达中的关键作用。我们进一步研究了METTL1/WDR4介导的tRNA m7G修饰是否可以调节胃癌细胞中的mRNA翻译。puromycin摄入分析显示METTL1的耗竭减少了胃癌细胞的整体mRNA翻译效率(TE)(图3J)。此外,野生型METTL1的重新表达(而非其突变体)在METTL1敲减的AGS和MGC803细胞中恢复了mRNA翻译效率(图3J),表明METTL1在mRNA翻译调节中的作用依赖于其甲基转移酶活性。这些发现强调了METTL1/WDR4介导的tRNA m7G修饰在调节胃癌中mRNA翻译的重要作用。
4)METTL1/WDR4以m7G相关密码子依赖的方式促进线粒体氧化磷酸化相关mRNA的翻译
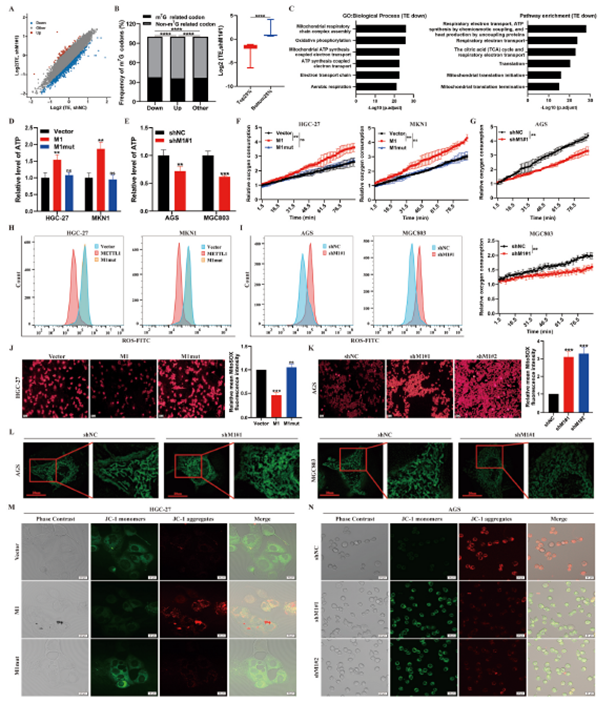
为了确定由METTL1/WDR4介导的m7G tRNA修饰调控的关键下游mRNA,我们在METTL1敲减和对照AGS细胞上进行了核糖体核酸复合物测序(RNC-seq),以识别翻译效率差异的mRNA。我们的分析发现,有871个mRNA的翻译效率降低(下调),而1179个mRNA的翻译效率增加(上调)(图4A)。随后,我们进行了密码子频率分析。与翻译效率增加的mRNA相比,翻译效率降低的mRNA具有更高比例的由m7G介导的tRNA解码的密码子(图4B)。接下来,我们使用RNC-seq数据进行了GO和通路富集分析。翻译效率低的mRNA在氧化磷酸化(OXPHOS)相关的生物学过程和与线粒体电子传递链(ETC)相关的通路中显著富集,这些过程和通路被认为是癌症进展的关键(图4C)。为了确认METTL1或WDR4的耗竭会破坏胃癌中的能量代谢,我们评估了ATP水平并进行了细胞外氧消耗(EOC)测定。野生型METTL1的过表达显著增强了胃癌细胞中的ATP合成和EOC,而METTL1的敲减则产生了相反的效果(图4D–G)。此外,METTL1或WDR4耗竭的胃癌细胞中的总细胞ROS产生和线粒体ROS水平显著上调,而在野生型METTL1或WDR4过表达时则下调(图4H,4J)。同时,MitoTracker Green染色实验显示,METTL1敲减导致AGS和MGC803细胞中线粒体形态异常,表现为线粒体变小、变圆、碎片化,正常的管状网络结构受损(图4L)。接下来,我们发现METTL1或WDR4的过表达会增加线粒体膜电位水平,而METTL1或WDR4的敲减则产生了相反的效果(图4M,4N)。总的来说,这些发现表明METTL1以一种依赖于m7G相关密码子的方式增强了与线粒体氧化磷酸化相关的基因的翻译。
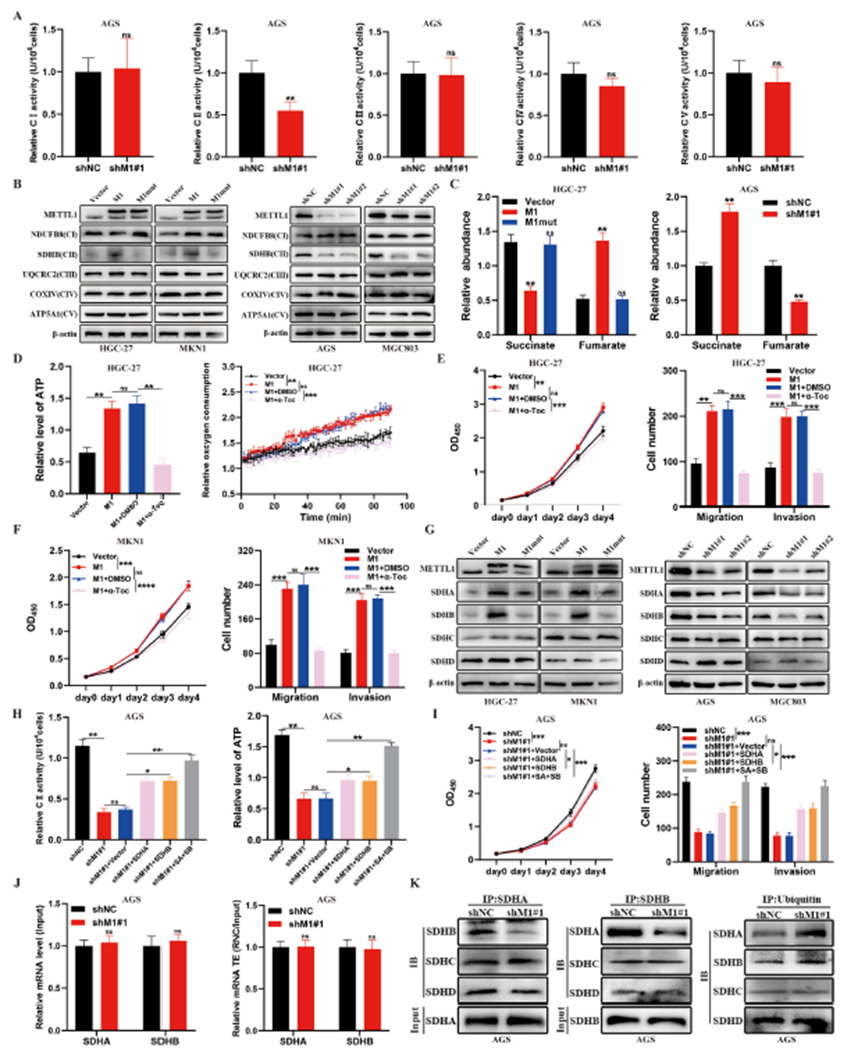
5)METTL1/WDR4介导的m7G tRNA修饰通过增强GC中ETC II活性促进氧化磷酸化和恶性进展
ETC对肿瘤生长至关重要。我们的RNC-seq结果显示,METTL1敲减影响了呼吸电子传递和ETC相关复合物的组装(图4C)。因此,我们推测METTL1通过调节ETC活性促进胃癌的OXPHOS和恶性进展。我们发现METTL1或WDR4敲减显著降低了ETC II的活性,而对其他复合物的活性没有影响(图5A)。此外,西方印迹分析显示,METTL1敲减降低了ETC II中的关键蛋白SDHB的表达水平,但对其他复合物中的关键蛋白表达没有影响(图5B)。相反,METTL1过表达增强了SDHB的表达(图5B)。METTL1敲减阻碍了延胡索酸的产生并导致琥珀酸的积累,而METTL1过表达则产生了相反的效果,进一步表明METTL1增强了ETC II的活性(图5C)。总的来说,这些发现表明METTL1可能增强了胃癌细胞中ETC II的活性。随后,我们使用ETC II抑制剂α-生育酚(α-Toc;50μM)来抑制METTL1过表达的HGC-27和MKN1细胞中ETC II活性。我们发现METTL1过表达会增加胃癌细胞中的ATP水平、EOC、增殖和转移能力,而ETC II抑制剂则产生了相反的效果。值得注意的是,用α-生育酚抑制ETC II的活性消除了METTL1过表达在胃癌细胞中引起的ATP水平、EOC、增殖和转移的增强效果(图5D–F)。这些结果证明了ETC II在介导METTL1在胃癌中的功能中的关键作用。
ETC II由四个亚单位蛋白组成(SDHA、SDHB、SDHC和SDHD)。METTL1影响了SDHA和SDHB的表达,但并未影响SDHC或SDHD(图5G)。我们在METTL1敲减的胃癌细胞中单独过表达SDHA和SDHB,以及共同过表达两者进行了拯救实验。SDHA或SDHB的重新表达部分恢复了METTL1敲减的胃癌细胞中的ETC II活性、ATP水平、增殖和转移能力,而同时过表达SDHA和SDHB则产生了更强的效果(图5H,5I)。此外,我们评估了METTL1敲减对SDHA和SDHB mRNA水平和翻译效率的影响。结果显示,METTL1敲减后,SDHA和SDHB mRNA的水平及其翻译效率没有显著变化(图5J),这表明SDHA和SDHB不是直接由METTL1介导的m7G tRNA修饰调控的。考虑到SDHA和SDHB蛋白表达减少的主要原因是泛素化水平增加,我们接着检查了METTL1敲减的胃癌细胞中SDHA和SDHB的泛素化水平。结果显示,METTL1敲减可以促进它们的泛素化水平,并减弱SDHA和SDHB之间的相互作用和结合(图5K和S6E)。总的来说,这些结果表明METTL1通过增强胃癌中依赖SDHA和SDHB的ETC II活性,促进氧化磷酸化和恶性进展。
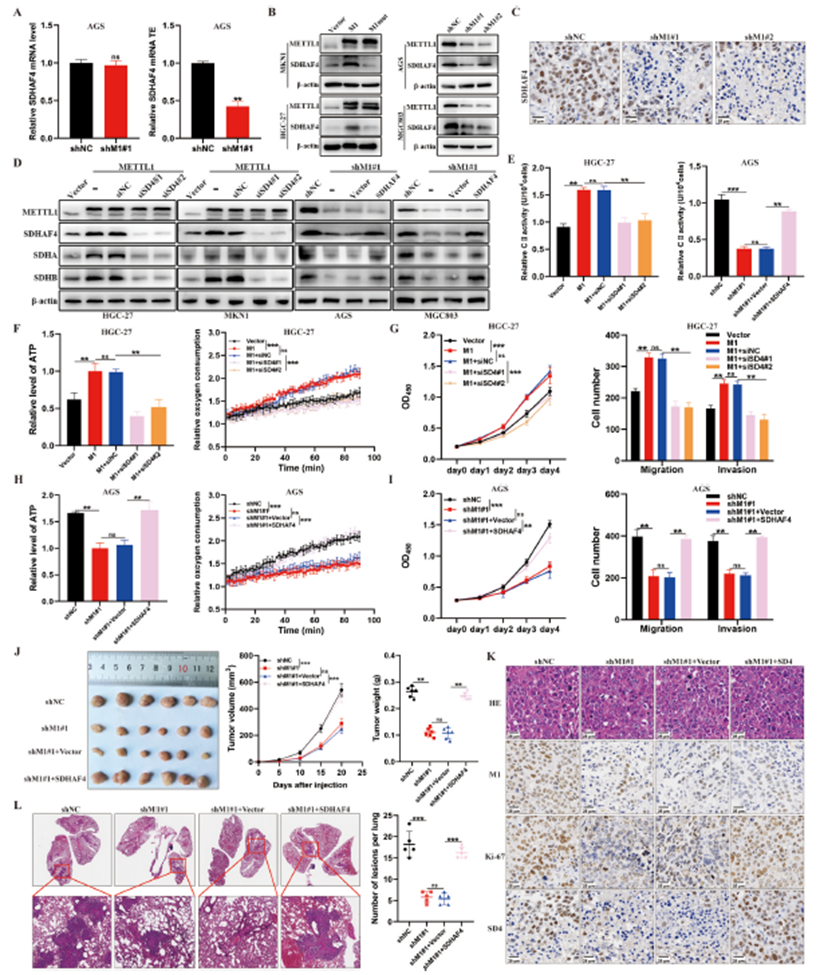
6)SDAHAF4是METTL1在GC中增强ETC II活性的关键靶点
鉴于METTL1并未影响胃癌细胞中SDHA或SDHB的翻译效率,我们分析了RNC-seq数据,以确定由METTL1介导的m7G tRNA修饰直接调控的、与ETC II活性相关的关键靶基因。我们发现,参与细胞呼吸和线粒体ETC复合物II组装的SDHAF4基因含有高丰度的m7G tRNA解码密码子。因此,我们假设SDHAF4是METTL1在胃癌中增强ETC II活性的关键靶基因。SDHAF4的翻译效率在METTL1敲减的细胞中降低,而其mRNA水平保持不变,表明METTL1/WDR4通过m7G tRNA修饰调控SDHAF4的表达(图6A)。此外,METTL1或WDR4耗竭时,胃癌细胞和异种移植肿瘤中的SDHAF4蛋白水平降低(图6B,6C)为了加强我们的结论,即METTL1介导的m7G tRNA修饰通过调控SDHAF4的翻译和表达增强了胃癌中ETC II的活性和恶性进展,我们在METTL1过表达的HGC-27细胞中使用了针对SDHAF4的siRNA,并通过西方印迹定量蛋白水平(图6D)。SDHAF4的敲减显著抑制了METTL1过表达引起的ATP水平、EOC、ETC II活性和细胞增殖、迁移和侵袭能力的增加(图6E–G)。相应地,METTL1敲减的MKN1细胞中恢复SDHAF4表达挽救了这些因素(图6E,6H,6I)。在皮下异种移植模型中,接受MKN45-shM1#1细胞治疗的小鼠肿瘤进展比接受MKN45-shNC细胞治疗的小鼠慢。值得注意的是,SDHAF4的过表达减弱了METTL1敲减对肿瘤进展的抑制效果(图6J)。此外,METTL1敲减降低了Ki-67、SDHAF4、SDHA和SDHB的表达,而过表达SDHAF4则逆转了METTL1敲减对Ki-67、SDHA和SDHB表达的抑制效果(图6K)。同样,METTL1敲减组的肺转移显著减少,而在体内重新过表达SDHAF4部分恢复了这些特性(图6L)。总的来说,这些发现表明SDHAF4对于METTL1增强胃癌中依赖SDHA和SDHB的ETC II活性和恶性进展至关重要。
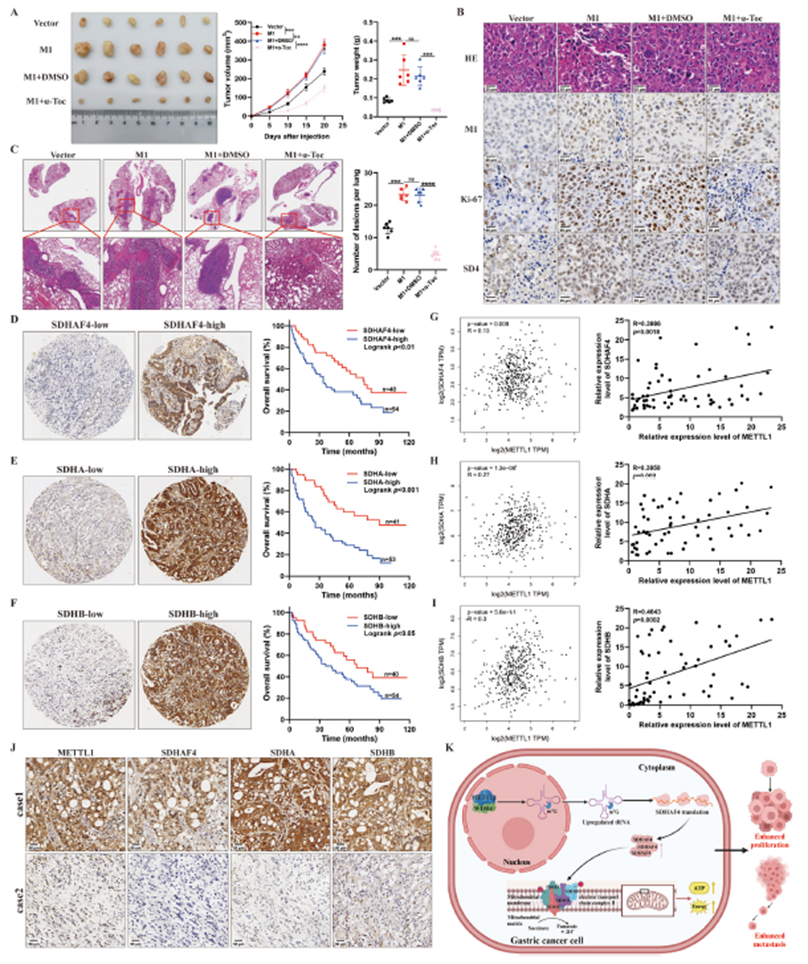
7)靶向METTL1/m7G/SDHAF4/SDHA/SDHB/ETC II轴可能是一种很有前景的GC治疗策略
最后,我们评估了针对METTL1/m7G/SDHAF4/SDHA/SDHB/ETC II轴的治疗是否可以作为胃癌的潜在治疗方法,以及这个轴是否对胃癌患者具有临床意义。METTL1的过表达显著加速了胃癌的进展,而α-生育酚对肿瘤生长表现出抑制效果,表明α-生育酚可能是高METTL1表达的胃癌患者的可行治疗选择(图7A)。免疫组化证实,METTL1过表达时Ki-67、SDHAF4、SDHA和SDHB的表达增加,而α-生育酚治疗逆转了这一效果(图7B)。在肺转移模型中,METTL1在体内显著促进了肺转移,而α-生育酚治疗减轻了这种效果(图7C)。Kaplan–Meier分析显示,SDHAF4、SDHA和SDHB水平与胃癌患者的预后不良呈正相关(图7D–F)。METTL1、SDHAF4、SDHA和SDHB的表达水平之间存在正相关(图7G–I)。此外,免疫组化染色验证了这一发现(图7J)。总的来说, METTL1/m7G/SDHAF4/SDHA/SDHB/ETC II轴在胃癌生长和转移中广泛参与,这表明针对这个轴的治疗可能作为胃癌治疗的充满希望的治疗策略。
结论:
我们的研究阐明了METTL1/WDR4介导的m7G tRNA修饰在胃癌进展中的致癌作用。此外,我们通过综合的体外、体内和临床样本数据揭示了通过m7G tRNA修饰在胃癌中调控特定mRNA转录本的分子机制。这些发现为针对胃癌开发治疗策略提供了理论基础。
实验方法:
qPCR,Northern,northwestern,western blotting,IHC,TRAC-seq,RNC- seq。
参考文献:
Xu X, Huang Z, Han H, Yu Z, Ye L, Zhao Z, Qian Y, Li Y, Zhao R, Zhang T, Liu Y, Cai J, Lin S, Zhai E, Chen J, Cai S. N7-methylguanosine tRNA modification promotes gastric cancer progression by activating SDHAF4-dependent mitochondrial oxidative phosphorylation. Cancer Lett. 2025 Apr 10;615:217566. doi: 10.1016/j.canlet.2025.217566.