






原来lncRNA阻碍侵袭也不一定是好事!
Pumilio(PUM)蛋白是高度保守的RNA结合蛋白家族的成员,该家族在转录后调节许多生物体中的基因表达。但是,它们在胎盘中的作用尚不清楚。胚胎着床时,滋养细胞粘附在母体子宫内膜上,然后侵入母体子宫内膜,最终形成胎盘。目前,在人类胎盘发育过程中发现了两种主要的滋养细胞谱系:绒毛滋养细胞和外渗滋养细胞(EVTs)。EVT的侵袭是胚胎植入和胎盘形成的重要过程。EVT侵袭不充分和子宫螺旋动脉重构受损是子痫前期(PE)发生的启动因素。本文确定了一种新的RNA调控机制,揭示了在PE发病机理中,调控PUM1 / HOTAIR在滋养细胞入侵中调控的新途径,并于今年12月发表在Molecular Therapy (IF: 8.402)。
主要结果如下:
1、PE患者滋养细胞中PUM1表达水平升高
收集妊娠中期绒毛组织,探讨PUM1和PUM2在PE发病中的作用,PUM1在PE中的mRNA和蛋白水平均上调(Fig 1A-B)。此外,与HC组相比,早发型PE组和晚发型PE组的绒毛组织中PUM1阳性信号更强(Fig 1C-F)。总之,PUM1在PE患者滋养细胞中高表达。
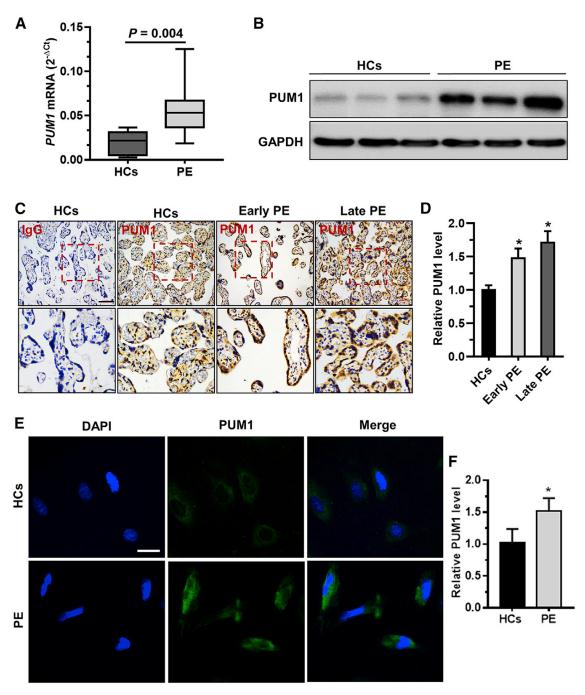
图1 PE患者滋养细胞中PUM1表达水平升高
2、PUM1抑制HTR-8细胞的增殖和侵袭
如图2所示,与对照组相比,干扰PUM1后HTR-8细胞的增殖和侵袭能力显著升高,表明PUM1抑制HTR-8细胞的增殖和侵袭。
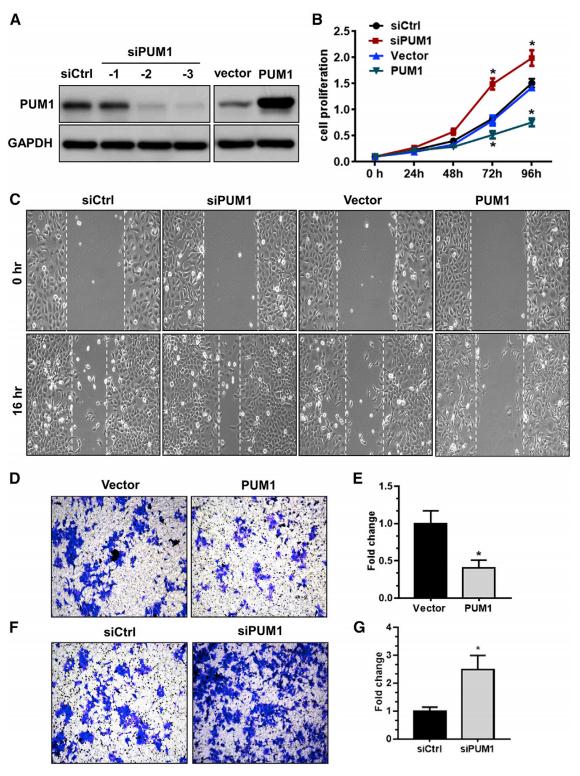
图2 PUM1抑制HTR-8细胞的增殖和侵袭
3、在绒毛外滋养细胞外植体培养模型中PUM1弱化了EVTs的入侵和迁移能力
为进一步研究PUM1在滋养细胞侵袭中的作用,从新鲜妊娠早期胎盘组织中分离出细胞滋养细胞(CT)和侵袭性滋养细胞(EVT)。采用qRT-PCR检测妊娠早期胎盘绒毛组织及CT、EVT细胞的PUM1 mRNA表达水平,结果发现,EVT细胞中PUM1 mRNA的表达水平低于CT细胞,表明PUM1可能在体内调控EVT的侵袭中发挥重要作用。
然后研究PUM1是否对滋养细胞的体外迁移能力有影响。从妊娠前3个月(妊娠6-10周)的胎盘中取出胎盘绒毛,分成两组,在涂有基质的培养皿中培养。培养24h后,siCtrl和siPUM1组之间没有观察到显着差异。体外培养72h后,siPUM1处理的外植体比siCtrl处理的外植体迁移得更远(图3A和3B)。全量免疫荧光染色显示了胎盘绒毛中PUM1的沉默效率(图3C和3D)。
为了进一步阐明PUM1对滋养层细胞入侵的调控,从健康的绒毛中获得新鲜外植体,并将其分为两组。结果表明,lenti-PUM1处理的外植体的迁移能力低于lenti-ctrl处理的外植体(图3E和3F)。综上,PUM1抑制EVT入侵并支持PUM1可能是PE的关键介体的观点。
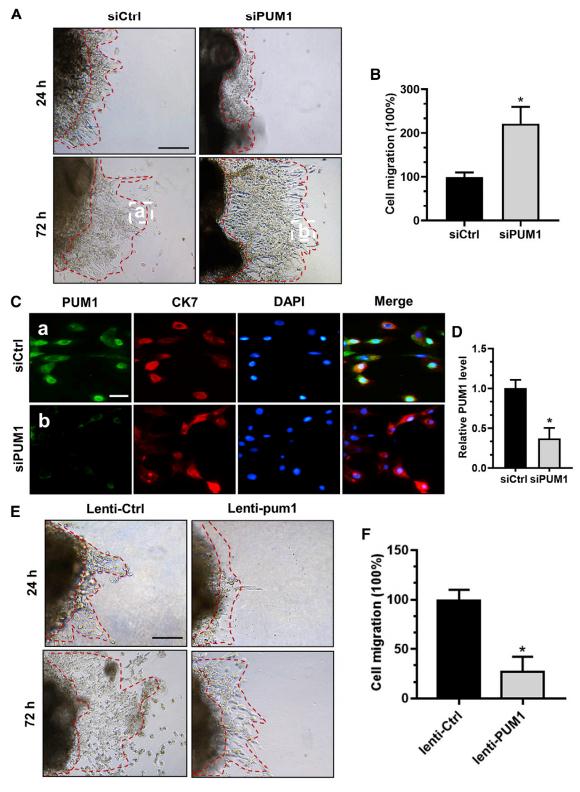
图3 PUM1减少了外植体中滋养层的生长
4、HOTAIR是PUM1在滋养层中的一个下游基因
为了阐明PUM1在PE发病机制中调控滋养细胞侵袭的分子机制,将HTR-8细胞转染siCtrl或siPUM1后培养48h的细胞进行lncRNA测序,得到178个差异表达lncRNA,与对照相比,siPUM1中101个上调77个下调(图4A);其中22个lncRNA的表达水平发生了显著变化 (图4B),经qRT-PCR验证,有12个的表达模式一直(图4C)。用PUM1过表达质粒转染HTR-8细胞,qRT-PCR结果显示,lncRNAs HOTAIR、HAGLR和CTD-2228K2.7在PUM1过表达组滋养细胞中表达较低;与此相反,MEG3、SNHG1、SNHG6和linc01085在PUM1过表达的滋养细胞中显著升高(图4D)。之后,使用RIP实验,结果发现只有HOTAIR显著富集(图4E)。此外,敲低PUM1可显著促进HOTAIR在HTR-8细胞中的表达,而过表达PUM1可降低HOTAIR的表达(图4F和4G)。总之,PUM1在胞质内与HOTAIR特异性相互作用,调节HOTAIR的表达。
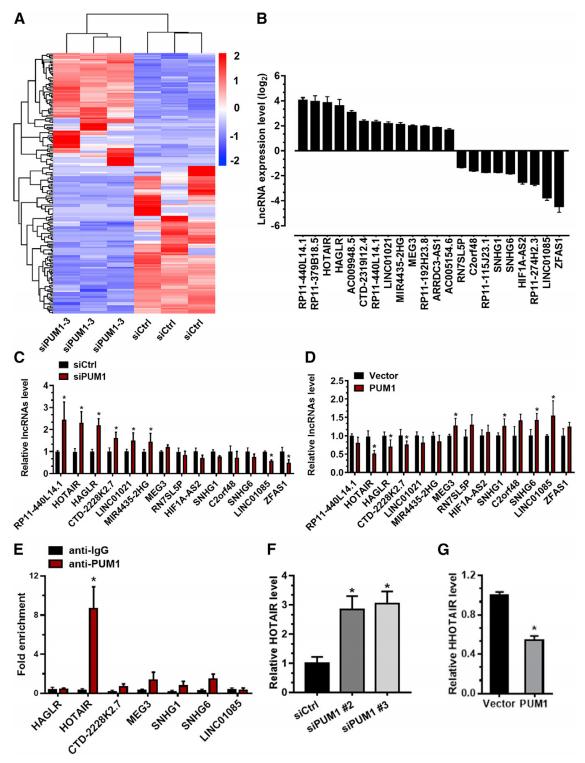
图4 PUM1下调滋养细胞中HOTAIR的表达
5、PUM1在滋养细胞中调节HOTAIR的表达和稳定性
为了进一步阐明PUM1调节滋养细胞中HOTAIR表达的潜在分子机制,根据UGUARAUA共有序列,在HOTAIR的第六个外显子区域(0.5 2.3 kb)中预测了四个PUM1结合位点,这些PUM1结合位点称为A,B,C和D(图5A)。荧光素酶结果显示,在对照组和PUM1沉默下,HTR-8细胞中MUT C的荧光素酶活性,特别是MUT D报告质粒的荧光素酶活性显着增加(图5B和5C),表明HOTAIR第六外显子区域中的位点C和D是PUM1的潜在结合位点。RIP结果显示,与IgG组相比,HOTAIR的C和D位点显着降低了抗PUM1组的HOTAIR富集(图5D),这些表明,HOTAIR的C和D位是结合PUM1蛋白的主要位点。RNA亲和纯化实验证实PUM1与HOTAIR特异性相互作用(图5E)。此外,转染siPUM1后,HOTAIR的半衰期较长,但当用过表达PUM1的质粒转染细胞时,HOTAIR的半衰期较短(图5F和5G),揭示了PUM1对HOTAIR RNA稳定性的强烈影响。综上,PUM1在转录后调控HOTAIR。
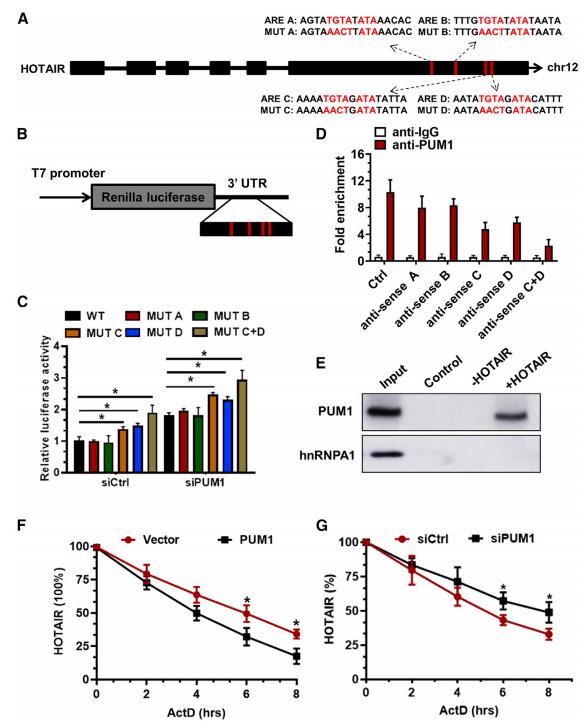
图5 PUM1是HOTAIR滋养细胞中重要的RNA结合蛋白
6、HOTAIR在PE中下调表达并与PUM1表达呈负相关
作者此前发现,HOTAIR通过激活PI3K- AKT信号通路来促进滋养细胞侵袭。实验发现,PUM1敲低或HOTAIR过表达增强了Ser473的AKT磷酸化,而PUM1过表达或HOTAIR敲低大大降低了AKT的磷酸化。此外,用HOTAIR过表达加PUM1过表达处理HTR-8细胞可以逆转PUM1过表达对AKT磷酸化的影响。相反,用HOTAIR siRNA加PUM1 siRNA处理HTR-8细胞也明显逆转了HOTAIR敲低对AKT磷酸化的作用(图6A和6B)。
此外,与对照组相比,过表达PUM1的HTR-8细胞和原代滋养细胞中侵袭细胞的数量明显减少,而HOTAIR的过表达逆转了由PUM1过表达引起的对滋养细胞入侵的影响(图6C-6F)。进一步验证PE患者和HCs中滋养细胞中HOTAIR和PUM1 mRNA的表达水平,和预期一致,与HC组相比,PE组的滋养细胞中PUM1 mRNA的水平明显较高,而HOTAIR的水平显着较低(图6G和6H)。线性相关分析表明,绒毛组织中HOTAIR水平与PUM1 mRNA水平呈负相关(图6I)。综上所述,PUM1 / HOTAIR / p-AKT途径在调节滋养细胞入侵中具有重要的功能。
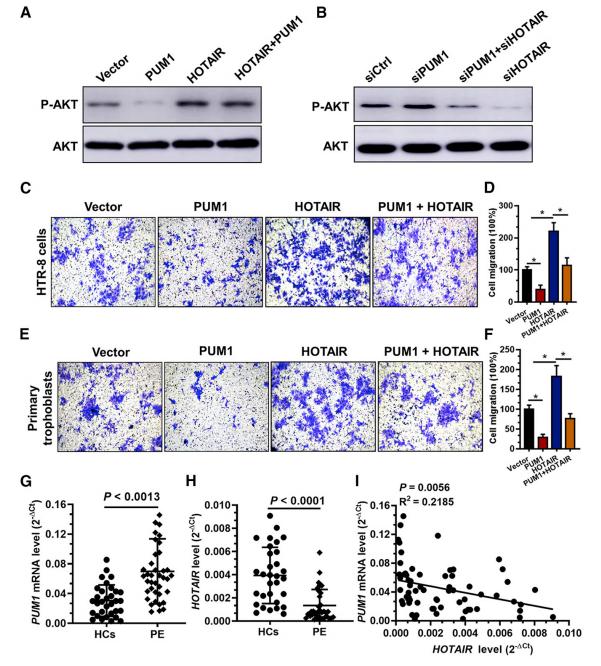
图6 PUM1/HOTAIR/AKT通路参与调节滋养细胞的侵袭
总之,本文表明PUM1是HOTAIR表达和滋养细胞功能的关键调控因子。首先,在PE中,PUM1上调,HOTAIR下调。其次,HOTAIR是滋养层迁移和入侵的有效调节因子,而在PE患者中,该过程不受控制。最后,AKT磷酸化的下调伴随着PUM1过表达后迁移和侵袭的减少。