






神经胶质细胞源性外泌体miR-274靶向触发气管和突触上按钮以调节生长和对缺氧的反应
神经胶质细胞源性外泌体miRNA通过下调基因表达来调节器官/组织之间的通讯,从而调节发育和生理功能。但是,尚未正式确定特定miRNA的来源,途径和功能。在这里,将显示神经胶质细胞源性外泌体miR-274非细胞自主调节突触钮扣和气管分支的生长。
技术路线:
饲养第三期幼虫用于实验cMyc及其调节剂在耐药中的作用
免疫染色,显微镜和图像处理解剖的三期幼虫
解剖游走的幼虫以进行EDC固定的荧光原位杂交(FISH)并检测前体miR-274
果蝇S2细胞培养物中分离外泌体并进行蛋白质印迹
从幼虫或外泌体组分中提取总RNA进行RT-PCR和实时PCR
游荡雌性幼虫RNA测序以及萤光素酶的构建和测定
对早期至三分龄幼虫的行为分析
使用Graphpad Prism v6进行统计分析
研究结果:
1.神经胶质细胞需要miR-274来调节突触和气管生长
通过透射电子显微镜进行的超微结构分析表明神经胶质-突触-气管组织可能代表功能单元,其形成可能受到发育调控。筛选了一组miRNA基因敲除突变体,定量分析证实了miR-274活性的缺乏是两个系统中生长缺陷的原因。同样也证明缺乏miR-274的幼虫无法发育出完整的突触按钮和气管分支。采用了由细胞类型特异性GAL4驱动程序驱动的UAS-decoy-mir-274转基因来抑制miR-274功能,证明了对miR-274的神经胶质抑制足以损害突触和气管生长。此外进行了神经胶质抢救实验,结果证明了神经胶质细胞表达的miR-274促进突触钮扣和气管分支的生长。
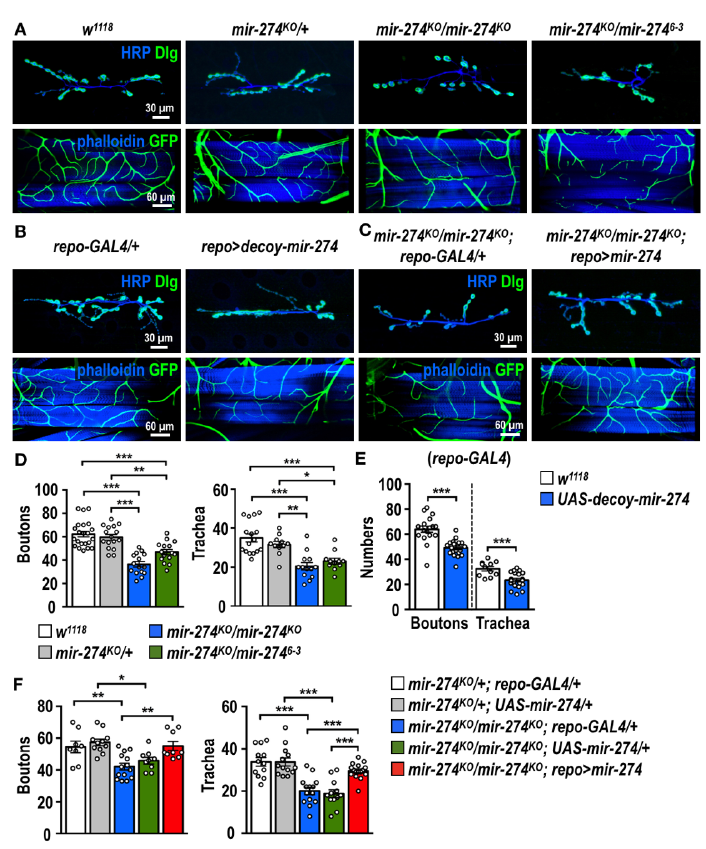
2.miR-274前体和成熟形式的表达
FISH分别检测了解剖的幼虫切片中的miR-274的前体或成熟形式。干扰探针在幼虫大脑中检测到低背景或非特异性信号。用于检测miR-274前体的环形探针在大脑中显示出明显的信号。这些信号位于由repo-GAL4驱动的mCD8-GFP标记的胶质细胞中,偶尔检测到强核信号。相反,在突触钮扣和气管分支中检测到低背景信号。使用干探针来检测成熟的miR-274。观察到了强大而普遍存在的信号。这些信号也在肌肉细胞和突触钮扣内以及气管体和分支中检测到。在mir-274KO突变体对照中未检测到成熟miR-274的信号。总结:miR-274前体主要在神经胶质中合成,并且成熟形式在肌肉,突触钮扣和气管细胞中检测到。
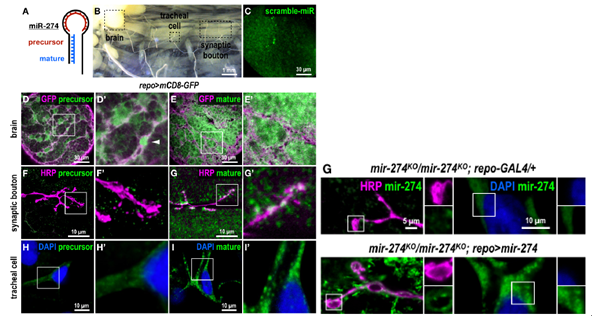
3.胶质细胞中miR-274的外泌体分泌
在S2细胞提取物中检测到miR-274。miR-274可能从S2细胞分泌到培养基中。S2细胞培养基分离的外泌体富含miR-274。在外泌体组分中检测到了外泌体标志物TSG101,Rab11和Syntaxin。在野生型幼虫的血淋巴中检测到大量的miR-274,而从mir-274KO突变体分离的血淋巴中则没有。在来源于野生型对照和突变幼虫的血淋巴的外泌体组分中检测到典型的外泌体标记物TSG101,Rab11和Syntaxin。因此,miR-274可以作为循环外泌体分泌到幼虫的血淋巴和S2细胞培养基中。检测神经胶质细胞可以在血液淋巴中分泌miR-274作为外泌体。我们对mir-274KO突变体中神经元和气管miR-274的表达进行了相同的实验。仅在整个幼虫裂解物中检测到miR-274,但在分离的血淋巴或外泌体组分中未检测到。通过绝对qPCR对血淋巴瘤中的miR-274水平进行定量。

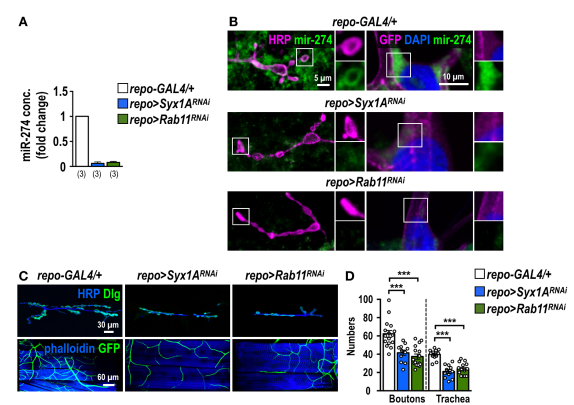
4.从胶质细胞中分泌miR-274外泌体需要ESCRT组件,Rab11和Syx1A
通过repo-GAL4在胶质细胞中表达了Rab11RNAi或Syx1ARNAi。从淋巴中分离出来的外泌体中miR-274转录水平显着降低。使用成熟的miR-274探针进行了FISH,,miR-274信号描绘了钮扣状的形态,并在肌肉和气管细胞中呈强烈的点状。在Rab11或Syx1A的胶质细胞敲低后未检测到这些miR-274信号。Rab11或Syx1A的胶质基因敲低导致突触钮扣和气管分支的生长减少。敲除ESCRT-I复合物的TSG101和ESCRT-III复合物的Shrb,胶质中TSG101或Shrb的RNAi敲除可有效抑制突触和气管生长。通过repo-GAL4在胶质细胞中特异性表达TSG101RNAi或shrbRNAi时,从淋巴液分离出的外泌体中miR-274的水平降低。这些结果强烈表明,miR-274是作为外泌体的货物分泌的,被释放到血淋巴中以调节突触和气管的生长。
5.Sprouty作为miR-274调控的靶基因
搜索具有miR-274靶位点并在mir-274KO幼虫中表达上调的基因。在最终的候选基因中,sty mRNA的3’UTR包含一个可识别miR-274的靶位点,具有靶向3'UTR的精确或错配的miR-274序列。当与miR-274共转染时, miR-274靶向序列下调了报道分子活性。sty mRNA的表达可能受miR-274通过其3'UTR靶向序列的调控。在mir-274KO幼虫中检测到了更高的sty转录水平。通过进行免疫染色,Sty受miR-274调节。检测突触终扣和野生型对照的气管分支的Sty的表达,Sty水平都得到了增强。这由Sty免疫荧光强度的定量支持。在肌肉细胞中,Sty表达也被上调,表明miR-274可能在多个组织中发挥系统调节作用。与野生型对照相比,mir-274KO的突触钮扣和气管细胞中dpERK的水平大大降低,dpERK水平的下调也取决于Sty。在肌肉中也检测到dpERK水平的恢复。减少miR-274突变体中的sty基因剂量可抑制两种生长表型。这些结果表明miR-274抑制Sty表达,从而导致MAPK活化以促进突触钮扣和气管分支的生长。

6.胶质细胞源性miR-274靶向突触钮和气管内的Sty,以调节其生长
首先在repo> decoy-mir-274幼虫中进行了免疫染色,该幼虫的突触和气管生长降低了。将miR-274捕获在胶质细胞中会导致突触后的bout和气管分支的Sty水平更高。检测到这两个位点的dpERK水平降低。发现胶质细胞拯救后,Sty降低,dpERK水平升高,并伴有突触钮扣数量增加和气管分支。这些结果说明胶质细胞表达的miR-274到达目标位点,以下调Sty表达并促进突触钮扣和气管分支的生长。最后,我们发现,该外来体生物发生,运输和释放由Rab11,Syx1A,TSG101和SHRB的胶质击倒的中断也引起麦粒肿上调和下调dpERK突触终扣和气管细胞。

7.miR-274调节幼虫缺氧反应
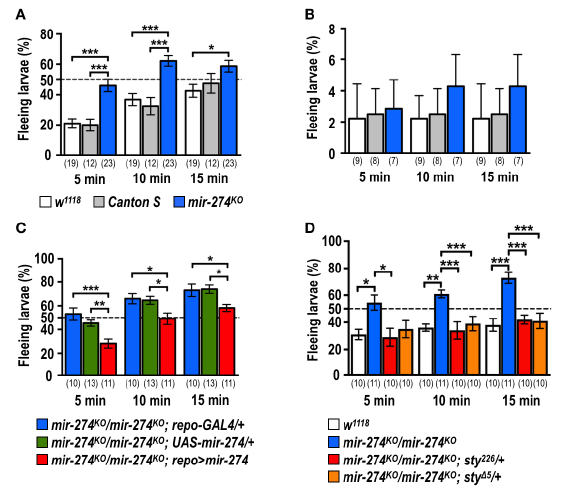
通过测定mir-274KO突变体的缺氧逃逸反应,当暴露于低氧环境中时,只有大约20%的对照幼虫在5分钟内通过逃离食物来源做出了强烈反应,到10分钟时,这一比例增加到几乎40%,接近15分钟后达到50%。相比之下,近50%的mir-274KO突变体在5分钟后表现出强烈的缺氧反应,在10和15分钟时表现出约60%的低氧反应。当我们在常氧环境下分析时,没有发现显着差异,几乎所有幼虫(> 95%)都留在食物中。此外,证实了低氧诱导的反应不是由于幼虫运动能力的差异所致,因为我们观察到了mir-274KO突变体和对照之间的爬虫长度相当。mir-274KO幼虫确实对较低的氧气含量具有更高的响应能力。进行了抢救实验,以检查正常幼虫缺氧反应是否需要胶质细胞表达的miR-274。与携带repo-GAL4或UAS-mir-274转基因的纯合子mir-274KO突变体相比,携带repo-GAL4和UAS-mir-274转基因的纯合子mir-274KO突变体显示出明显降低的缺氧反应。
通过突变进行了Sty的遗传抑制,发现在纯合的mir-274KO突变体中引入sty突变体等位基因几乎完全抑制了缺氧逃逸反应的增强。在不存在miR-274的情况下,sty突变等位基因也可恢复气管分支,因此该结果支持气管分支与缺氧逃逸反应相关。
总 结:
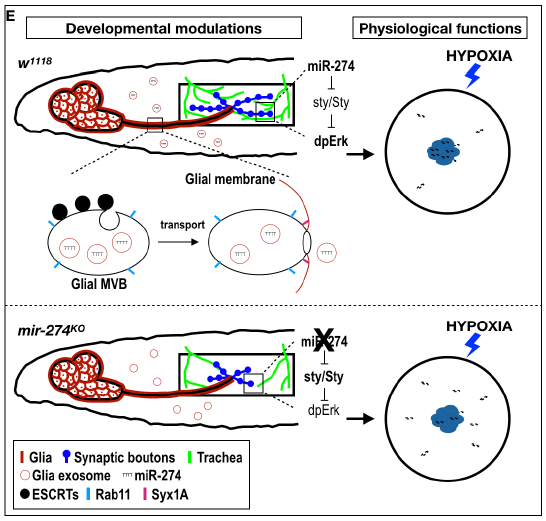
在这项研究中,作者探索了果蝇幼虫神经肌肉接头(NMJ),其中轴突末端分支形成带有肌肉膜的突触钮扣。我们证明NMJ还非常重视气管末端分支,使其成为研究神经和血管协调发育的理想系统。作者筛选了一组miRNA敲除突变体,并确定mir-274突变体在突触和气管生长中均具有缺陷。通过荧光原位杂交(FISH),表明miR-274前体在胶质细胞中表达,成熟形式被普遍检测。一致地,在胶质细胞中需要miR-274来进行突触和气管生长。可以在幼虫循环系统的血淋巴中检测到miR-274的神经胶质表达。确实,如遗传分析和生化分级分析所示,miR-274被分泌为外泌体货物。我们鉴定出一个miR-274靶标发芽(sty),该靶标在sty转录本的3'UTR中包含一个靶标位点,并表明神经胶质miR-274的表达可诱导突触管和气管分支中的Sty下调和MAPK活化。有趣的是,气管分支较少的mir-274突变体对缺氧过敏,并且两种表型均被抑制。