






单细胞转录组学阐明了嗜碱性粒细胞分化轨迹, 并鉴定了成熟嗜碱性粒细胞上游的前嗜碱性粒细胞
嗜碱性粒细胞是最稀有的粒细胞,被认为是2型免疫反应的关键细胞。然而,它们的分化路径仍未完全阐明。在这项研究中,我们通过单细胞RNA测序分析来评估嗜碱性粒细胞的个体发育轨迹。结合流式细胞术和功能分析,我们发现c-Kit-CLEC12Ahi前嗜碱性粒细胞位于前嗜碱性粒细胞和肥大细胞祖细胞(pre-BMPs)下游和CLEC12Alo成熟嗜碱性粒细胞上游。转录分析预测,在基因表达谱方面,前嗜碱性粒细胞群体包括先前定义的嗜碱性粒细胞祖(BaP)-like 细胞。与成熟嗜碱性粒细胞相比,前嗜碱性粒细胞具有较高的增殖能力,对非IgE刺激的反应较好,但对抗原加IgE刺激的反应较弱。尽管前嗜碱性粒细胞通常停留在骨髓中,但它们会在蛔虫感染的组织中出现,可能是通过IL-3介导抑制其在骨髓中的滞留。因此,本研究确定了在嗜碱性粒细胞个体发育过程中连接pre-BMPs和成熟嗜碱性粒细胞之间的前嗜碱性粒细胞。
该研究于2023年5月发表发表在《Nature communications》,IF:17.694。
技术路线:

实验方法:抗体、细胞培养、流式细胞仪分析与细胞分选、Giemsa染色、scRNA-seq、bulk RNA-seq分析、蠕虫感染、嗜碱性粒细胞分类纯化后的体外细胞培养、统计分析。
1、骨髓来源的嗜碱性粒细胞(BMBAs)包含两个亚群
嗜碱性粒细胞是数量最少、寿命较短的粒细胞,因此常常使用骨髓来源的嗜碱性粒细胞(BMBAs)作为新鲜动物嗜碱性粒细胞的替代物。为了确定BMBAs的最佳培养条件,我们将小鼠骨髓细胞与不同浓度的IL-3进行培养,浓度范围从0到100 ng/mL,培养时间为7天。随着IL-3浓度增加至0.3 ng/mL,培养物中CD200R3+ c-Kit−嗜碱性粒细胞的比例增加到约60%。在较高浓度的IL-3下,嗜碱性粒细胞的比例反而下降,而CD200R3+ c-Kit+肥大细胞的比例增加。因此,0.3 ng/mL的IL-3被证明是制备高纯度嗜碱性细胞的最佳选择,虽然远低于通常使用的浓度。
在上述实验中,我们注意到CD200R3+ c-Kit− BMBAs中存在两个亚群:FcεRIαhiCD49blo和FcεRIαloCD49bhi(图1a)。不论使用C57BL/6还是BALB/c小鼠,前者亚群的频率随着IL-3浓度增加而下降,表明IL-3对这两个亚群之间的平衡产生影响。值得注意的是,FcεRIαloCD49bhi BMBAs显示出与外周血嗜碱性粒细胞相同的环状细胞核,而FcεRIαhiCD49blo BMBAs则显示出非典型形态,呈肾形凹陷状,胞体较大(图1b)。FcεRIαhiCD49blo BMBas由于细胞体较大,其前向散射强度高于FcεRIαloCD49bhi BMBas(图1c)。
为了进一步表征和比较这两个BMBAs亚群,我们对骨髓细胞的IL-3培养物中的细胞进行了无偏向单细胞RNA测序(scRNA-seq)分析。通过Seurat聚类分析scRNA-seq数据集,确定了11个细胞簇,其中簇0、簇2和簇10是根据Cd200r3和Mcpt8的高表达以及Kit的低表达被判定为嗜碱性粒细胞。在这三个簇中(分别在图1d中表示为BMBA1、BMBA2和BMBA3),BMBA2和BMBA3的Fcer1a(编码FcεRIα)表达较低,而Itga2(编码CD49b)表达较高,相比之下BMBA1则相反(图1d、e)。因此,BMBA2和BMBA3似乎对应于FcεRIαloCD49bhi BMBAs,而BMBA1对应于FcεRIαhiCD49blo BMBAs。在BMBA1和BMBA2/3之间,我们检测到5382个差异表达基因(DEGs),其中4811个基因在BMBA1中上调,而571个基因在BMBA2/3中上调(图1f)。其中,我们选择了两个编码细胞表面蛋白的基因,Clec12a编码CLEC12A和Cd9编码CD9(图1e、f),作为可能的代用标记物,用于在后续实验中区分BMBAs的两个亚群。使用特异性单克隆抗体进行的流式细胞术分析显示,FcεRIαhiCD49blo和FcεRIαloCD49bhi BMBAs几乎对应于CLEC12AhiCD9lo和CLEC12AloCD9hi BMBAs(图1g)。同样,CLEC12AhiCD9lo和CLEC12AloCD9hi BMBAs几乎对应于FcεRIαhiCD49blo和FcεRIαloCD49bhi BMBAs(图1h)。
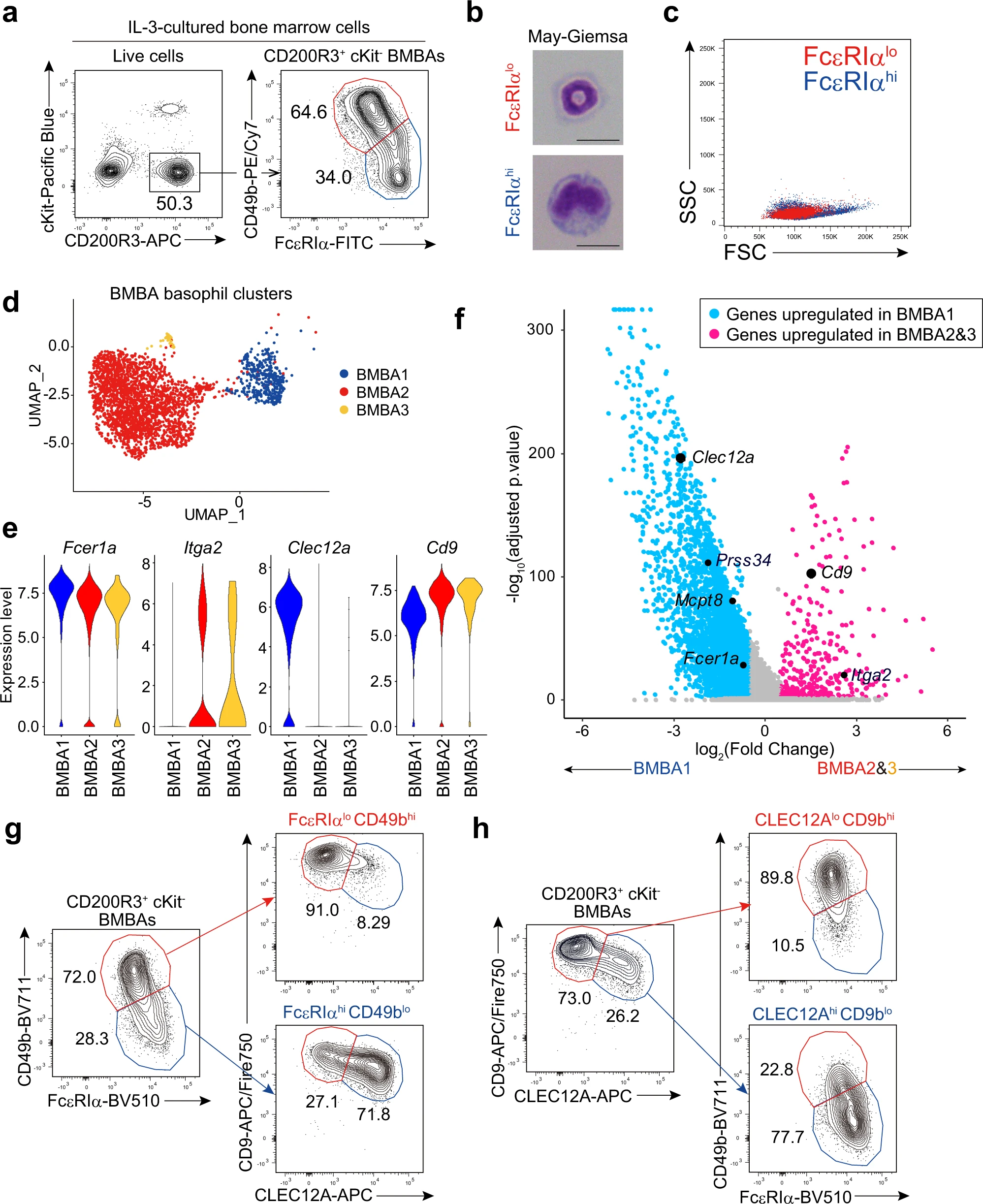
图1|骨髓源性嗜碱性粒细胞(BMBA)由两个不同的亚群组成。
2、在骨髓中检测到两个嗜碱性粒细胞的亚群,而在外周血和脾脏中只检测到其中的一个亚群
接下来,我们检查了是否可以在从小鼠中新鲜分离的嗜碱性粒细胞中检测到在BMBAs中鉴定出的两个嗜碱性粒细胞亚群。在c-Kit-CD200R3+ CD49b+嗜碱性粒细胞中,FcεRIαloCD49bhi/CLEC12AloCD9hi亚群在外周血、脾脏和骨髓中都很容易被鉴定出来(图2a)。相反,尽管骨髓中存在,但在外周血和脾脏中很少检测到FcεRIαhiCD49blo /CLEC12AhiCD9lo亚群(图2a)。骨髓、脾脏和外周血中的CLEC12AloCD9hi嗜碱性粒细胞具有典型的环形或双裂核,而骨髓中的CLEC12AhiCD9lo嗜碱性粒细胞则显示出具有肾脏形状的凹陷核和较大的细胞体,并在流式细胞术中显示出较高的前向散射(FSC),这与BMBAs中观察到的情况相符(图2b、c)。值得注意的是,Lin-cKit-CD34+ CD200R3+ BaPs也显示出CLEC12AhiCD9lo表型,而在cKit-CD200R3+嗜碱性粒细胞谱系细胞中,约10%的CLEC12Ahi亚群CD34表达量较低(图2d),这表明CLEC12Ahi嗜碱性粒细胞可能包含单能的类似BaP的群体。支持这一观点的是,CD34+ BaPs和CD34- CLEC12Ahi嗜碱性粒细胞显示出类似的形态和表面表达特征也支持了这一观点。

图2|在骨髓中也检测到两个亚群的嗜碱性粒细胞,而在外周血和脾中只检测到一个亚群
3、单细胞RNA测序(scRNA-seq)分析鉴定出骨髓嗜碱性粒细胞中的4个簇
我们使用Mcpt8GFP转基因小鼠进行分析,该转基因小鼠在Mcpt8启动子/增强子下表达GFP编码基因,以有效富集和分离稀有的嗜碱性粒细胞,以分析骨髓嗜碱性粒细胞的分化轨迹。我们确认,CLEC12Ahi和CLEC12Alo嗜碱性粒细胞亚群中的大多数细胞都表达GFP。从骨髓和脾脏中分离的Lineage(Lin)- GFP+细胞分别进行了单细胞RNA测序分析。单细胞RNA测序数据的聚类分析鉴定出12个簇,其中0、1和4号簇对应于KitCd200r3+ Mcpt8+嗜碱性粒细胞。对这三个簇进行重新聚类,鉴定出1个Fcer1a+ Kit+Cd34+类似于pre-BMPs细胞的簇,1个Clec12ahi嗜碱性粒细胞簇(Baso1)和2个Clec12alo嗜碱性粒细胞簇(Baso2和Baso3)(图2e、f)。与流式细胞术分析结果一致,Clec12ahi嗜碱性粒细胞(Baso1)在骨髓中丰富,而在脾脏中较少(图2e、f)。与流式细胞术分析一致,21%的Clec12ahi Baso1细胞的Cd34表达水平较低(图2f)。
为了验证我们的发现,我们重新分析了由Weinreb等人报道的公开可用的scRNA-seq数据集(GEO登录号:GSE140802),鉴定出1个Fcer1a+Kit+Cd34+类似于preBMPs细胞的簇,2个Clec12ahi嗜碱性粒细胞簇(Baso1和Baso2),以及3个Clec12alo嗜碱性粒细胞簇(Baso3、Baso4和Baso5)。根据我们的scRNAseq数据分析,仅有小部分(11.4%)的Clec12ahi嗜碱性粒细胞显示Cd34表达水平较低,这表明Clec12ahi嗜碱性粒细胞中的这一小部分可能对应于Cd34lo类似于BaP的细胞。
4、CLEC12Ahi嗜碱性粒细胞在体外和体内分化为CLEC12Alo嗜碱性粒细胞
对我们的scRNA-seq数据进行的伪时间轨迹分析和RNA速度分析推断嗜碱性粒细胞的分化轨迹,从前BMPs-like细胞到Clec12ahi嗜碱性粒细胞,然后到Clec12alo嗜碱性粒细胞 (图2g,h)。
为了验证假设,我们追踪了从骨髓中分离出,并在体外培养2天的c-Kit-嗜碱性粒细胞的FcεRIαhiCD49blo和FcεRIαloCD49bhi组分的表面CLEC12A和CD9表达的变化。在培养过程中,大多数的FcεRIαhiCD49blo嗜碱性粒细胞的表面表型从CLEC12AhiCD9lo转变为CLEC12AloCD9hi(图3a,左图)。相反,FcεRIαloCD49bhi嗜碱性粒细胞即使在培养2天后仍然保持CLEC12AloCD9hi表型(图3a,右图)。值得注意的是,FcεRIαloCD49bhi 嗜碱性粒细胞在2天的培养过程中细胞总数减少了约90%,而FcεRIαhiCD49blo 嗜碱性粒细胞的减少仅约为40%(图3b,c),这表明前者的寿命较短。当我们分别培养骨髓嗜碱性粒细胞的CLEC12AhiCD9lo和CLEC12AloCD9hi亚群,并追踪FcεRIα和CD49b表面表达的变化时,也获得了相符的结果。这些结果强烈表明,CLEC12AhiCD9lo/FcεRIαhiCD49blo 嗜碱性粒细胞相对不成熟,并且可以分化为CLEC12AloCD9hi/FcεRIαloCD49bhi成熟型嗜碱性粒细胞,尽管后者的Fcer1a、Mcpt8和Prss34表达水平低于前者(图2i)。
接下来,我们检查了体外实验的体内相关性。FcεRIαhiCD49blo和FcεRIαloCD49bhi组分分别从CD45.2+小鼠的骨髓细胞中分类纯化并过继转移到亚致死剂量照射的CD45.1+小鼠(图3d)。与体外实验观察到的结果一致,大多数的FcεRIαhiCD49blo 嗜碱性粒细胞在骨髓中将它们的表面表型从CLEC12AhiCD9lo转变为CLEC12AloCD9hi,而大多数的FcεRIαloCD49bhi 嗜碱性粒细胞仍保持CLEC12AloCD9hi(图3d)。
综上所述,我们得出结论,骨髓中的CLEC12AhiCD9lo/ FcεRIαhiCD49blo 嗜碱性粒细胞在表面标记物的表达、细胞大小和核形状上显示出前体like的表型,并在骨髓内分化为CLEC12AloCD9hi/ FcεRIαloCD49bhi成熟嗜碱性粒细胞,这些嗜碱性粒细胞也存在于外周血和脾脏中。因此,在接下来的实验中,我们将CLEC12AhiCD9lo/ FcεRIαhiCD49blo和CLEC12AloCD9hi/FcεRIαloCD49bhi 嗜碱性粒细胞分别称为前体嗜碱性粒细胞和成熟嗜碱性粒细胞。
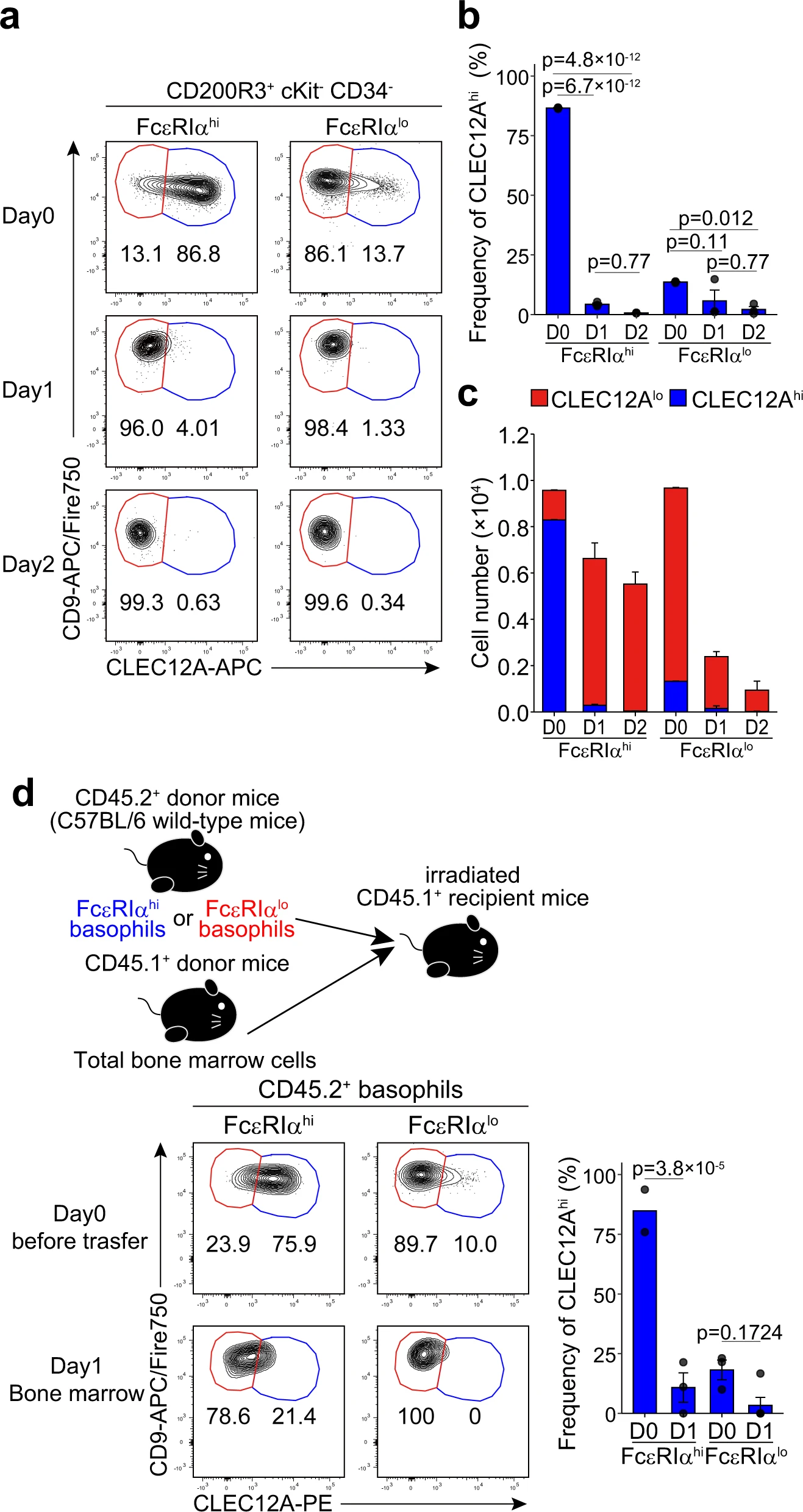
图3| CLEC12Ahi嗜碱性粒细胞在体外和体内分化为CLEC12Alo嗜碱性粒细胞
5、前嗜碱性粒细胞和成熟嗜碱性粒细胞表现出不同的细胞增殖和激活特性
为了进一步鉴定前嗜碱性粒细胞(pre-嗜碱性粒细胞)和成熟嗜碱性粒细胞(mature 嗜碱性粒细胞),我们分别从骨髓中分离了CLEC12AhiCD9loCD34-的前嗜碱性粒细胞以及从骨髓和脾脏分离的CLEC12AloCD9hi的成熟嗜碱性粒细胞,并利用批量RNA测序分析比较它们的基因表达谱(图4a)。作为参考,我们还包括了纯化的CD34+ BaPs,它们被定义为单功能嗜碱性粒细胞前体细胞。DEGS的系统聚类分析表明,从骨髓和脾脏分离的成熟嗜碱性粒细胞的基因表达谱彼此相似,而CD34-的前嗜碱性粒细胞和成熟嗜碱性粒细胞的基因表达谱显示出明显差异(图4a)。值得注意的是,CD34-的前嗜碱性粒细胞的基因表达谱与CD34+BaPs的基因表达谱相似(图4a),加强了我们之前关于CD34+BaPs被包括在CLEC12Ahi前嗜碱性粒细胞群体中的推测。
GO富集分析显示,与细胞增殖相关的GO术语,如“核分裂”、“染色体分离”和“DNA复制”,在CLEC12Ahi前嗜碱性粒细胞中富集,与CLEC12Alo成熟嗜碱性粒细胞相比(图4b),单细胞RNA测序分析结果显示,与CLEC12Alo Baso2/3群体相比CLEC12Ahi Baso1群体显示出较高的S期得分。实际上,在前嗜碱性粒细胞中明显检测到EdU核苷类似物的摄取,而成熟嗜碱性粒细胞中未检测到(图4d),这表明前嗜碱性粒细胞的增殖能力。前嗜碱性粒细胞和成熟嗜碱性粒细胞在增殖能力上的差异似乎可以解释在2天培养过程中观察到的细胞总数减少的差异(图3c)
与前嗜碱性粒细胞相比,成熟嗜碱性粒细胞富集了与免疫效应功能相关的GO术语,如“调节免疫效应过程”和“正调节细胞因子产生”(图4c)。当受到IgE加抗原的刺激时,CLEC12Alo成熟嗜碱性粒细胞显示出比CLEC12Ahi前嗜碱性粒细胞更高水平的CD63表达(代表颗粒脱颗粒的指标)和IL-4产生(图4e,f),尽管成熟嗜碱性粒细胞表面FcεRI表达水平较低(图2a)。有趣的是,当受到IL-3、IL-33和LPS等先天型刺激时,前嗜碱性粒细胞产生比成熟嗜碱性粒细胞更高水平的IL-4(图4g)。此外,批量RNA测序分析发现,在IL-3刺激的前嗜碱性粒细胞中,其基因表达谱与IL-3或抗原/IgE刺激的成熟嗜碱性粒细胞有所不同,包括前者中Il10和Il13的上调表达。因此,前嗜碱性粒细胞似乎对非IgE刺激更具反应性,而成熟嗜碱性粒细胞对IgE/变应原刺激更具反应性。
根据细胞因子环境的不同即在嗜碱性细胞产生过程中是否存在IL-3或TSLP ,两种具有不同表型和功能特性的嗜碱性粒细胞群体被报道。当受到IL-3和IL-33的刺激时,TSLP诱导的嗜碱性粒细胞比IL-3诱导的嗜碱性粒细胞产生更多的IL-4和IL-6。相反,在脱颗粒方面,TSLP诱导的嗜碱性粒细胞对IgE交联的反应较差。这种TSLP诱导的嗜碱性粒细胞的反应性似乎与本研究中鉴定的前嗜碱性粒细胞相似。然而,TSLP诱导的嗜碱性粒细胞在大小上相对较小,并具有成熟型嗜碱性粒细胞特有的环状核。据此,我们发现大多数TSLP诱导的嗜碱性粒细胞表现为成熟嗜碱性粒细胞(FcεRIαloCD49bhi/CLEC12AloCD9hi表型),而不是前嗜碱性粒细胞。此外,TSLP诱导的成熟嗜碱性粒细胞显示出类似于IL-3诱导的成熟嗜碱性粒细胞的基因表达谱,而不是前嗜碱性粒细胞。
综上所述,TSLP诱导的嗜碱性粒细胞似乎不对应于前嗜碱性粒细胞,可被归类为成熟嗜碱性粒细胞的一个亚群。
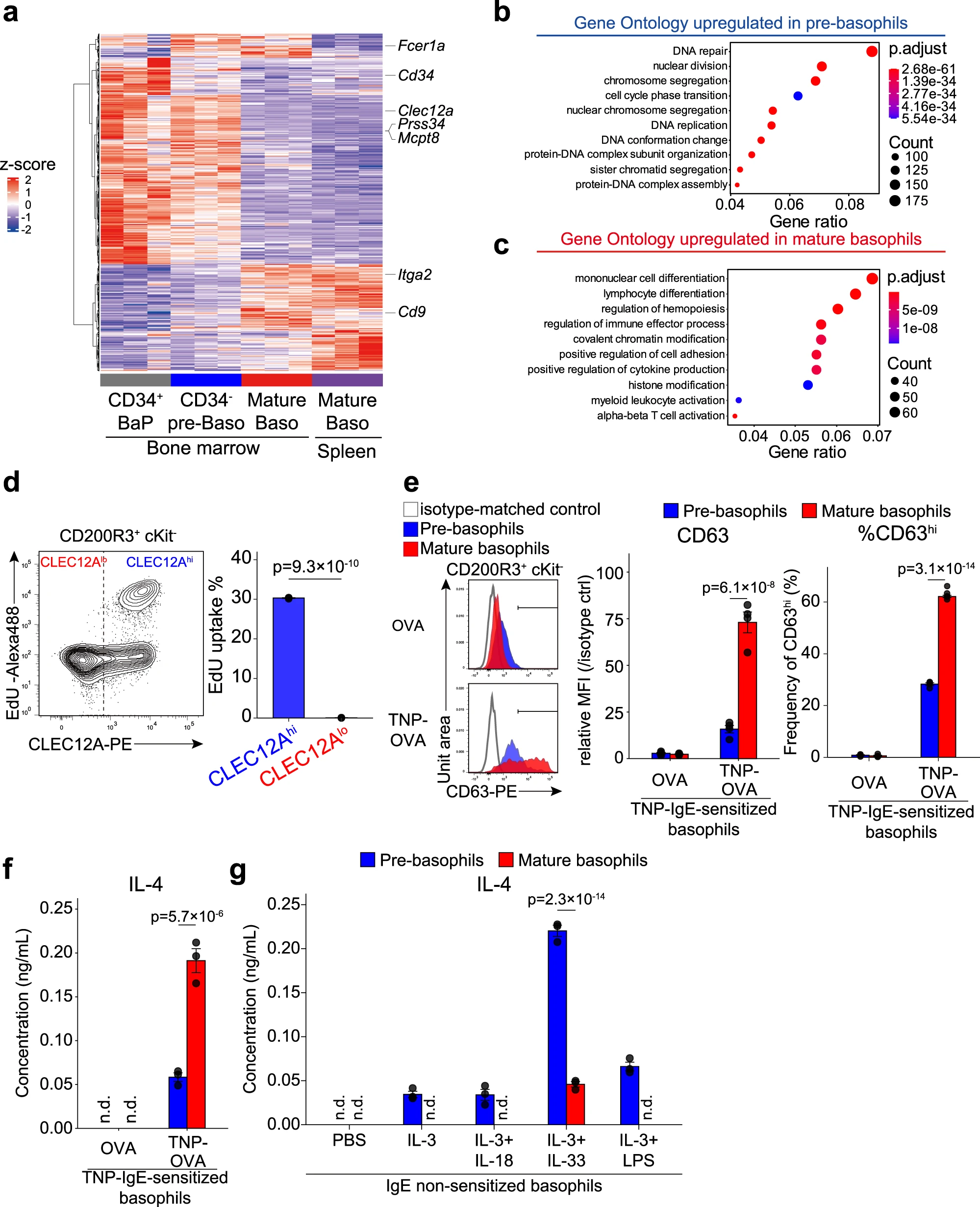
图4|前嗜碱性粒细胞和成熟嗜碱性粒细胞表现出不同的细胞增殖和激活特性
6、在蠕虫感染期间,周围组织中不仅出现成熟的嗜碱性粒细胞,而且还出现前嗜碱性粒细胞
在寄生虫感染期间,嗜碱性粒细胞常常浸润到外周组织中。巴西尼普斯壮线虫(Nb)是一种在人类钩虫感染的小鼠模型中被广泛研究的蠕虫。它们通过皮肤进入宿主动物,迁移到肺部,最终到达肠道。寄生虫通过肺部后,嗜碱性粒细胞在肺部积累,并且对抑制过度肺部炎症和修复受损肺组织至关重要。在Nb感染后的第7天,我们在脾脏、血液甚至肺部除了CLEC12Alo成熟嗜碱性粒细胞外,还检测到CLEC12Ahi前嗜碱性粒细胞(图5a),这与未感染或PBS注射对照小鼠中前嗜碱性粒细胞主要存在于骨髓中的观察结果相反(图2a)。从Nb感染的肺部分离的前嗜碱性粒细胞显示出比成熟嗜碱性粒细胞更高的体外EdU掺入(图5b),这表明前嗜碱性粒细胞即使在骨髓之外仍保持着较高的增殖能力。此外,在感染的肺部积累的前嗜碱性粒细胞与成熟嗜碱性粒细胞的IL-4表达相当,这暗示它们可能对Nb感染的Th2免疫起到贡献作用(图5c)。
我们以前曾证明在二次Nb感染中,嗜碱性粒细胞渗入受感染的皮肤,并将蠕虫困在皮肤内,以防止感染进一步传播到肺部和肠道。在第二次NB感染的皮损中,除了CLEC12Alo成熟的嗜碱性粒细胞外,还检测到了前嗜碱性粒细胞(图5d)。从骨髓和第二次感染的皮肤中分离的CD200R3+ cKit-细胞的单细胞RNA测序分析鉴定了9个聚类,其中聚类0、1和2对应于Cd200r3+ Mcpt8+嗜碱性粒细胞。在这三个嗜碱性粒细胞聚类中,我们在两个器官中都检测到一个Clec12ahi前嗜碱性粒细胞聚类和两个Clec12alo成熟嗜碱性粒细胞聚类(图5e,f)。对来自Nb感染和未感染小鼠的嗜碱性粒细胞的单细胞RNA测序数据进行整合分析发现,在Nb感染小鼠的皮肤中检测到的前嗜碱性粒细胞的基因表达谱与未感染小鼠骨髓中的前嗜碱性粒细胞非常接近,包括Fcer1a,Clec12a,Mcpt8和Prss34的上调表达(图5f)。此外,基因集富集分析(GSEA)和GO富集分析发现,在Nb感染的皮肤中,前嗜碱性粒细胞富集了与细胞增殖相关的基因,包括“染色体分离”(图5g),这表明前嗜碱性粒细胞在感染的皮肤中保持着其高增殖能力,在感染的肺部观察到的结果也一样。
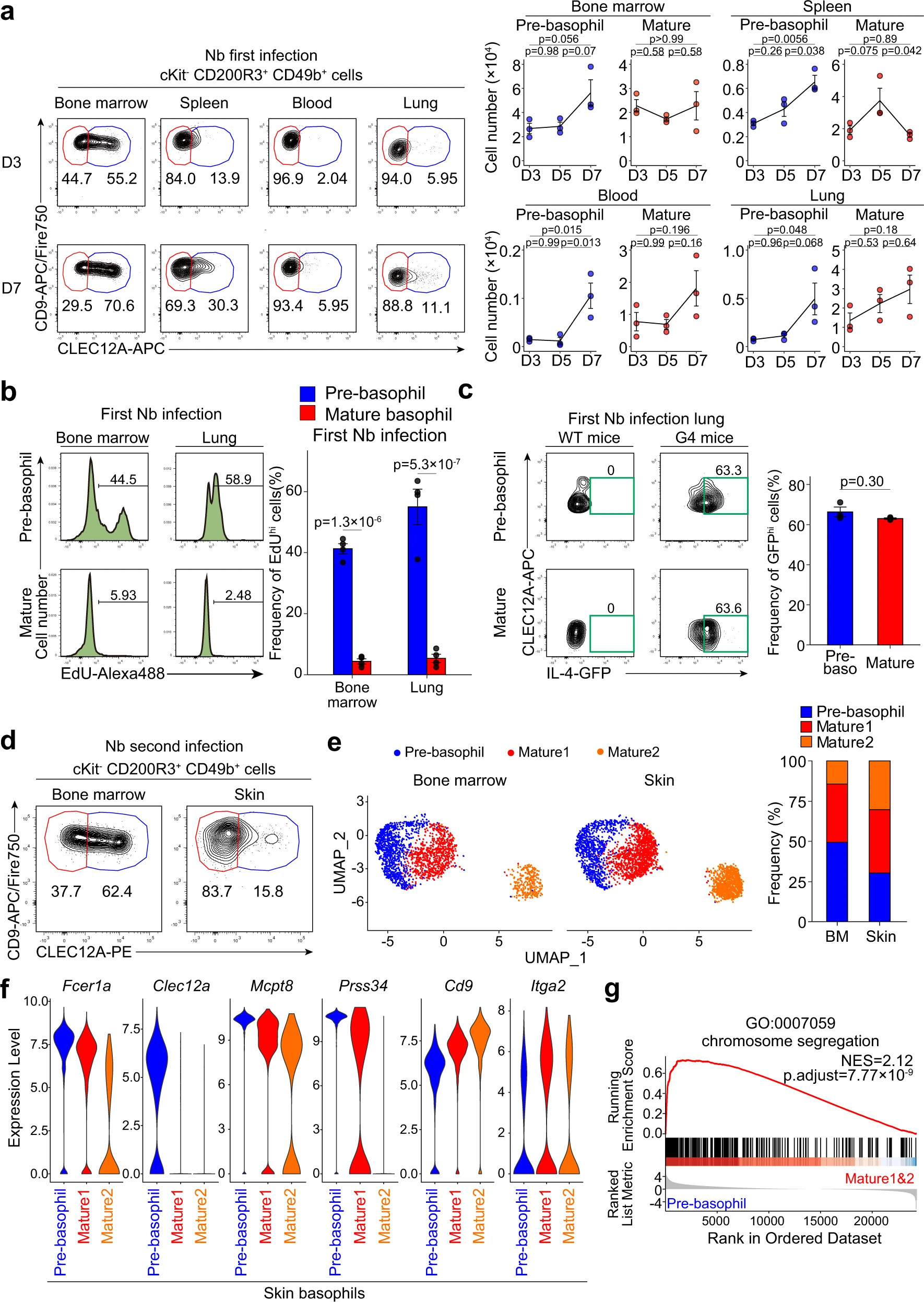
图5|蠕虫感染期间外周组织中可检测到前嗜碱性粒细胞
7、前嗜碱性粒细胞中IL-3介导的CXCR4下调促进其从骨髓中排出
接下来,我们试图阐明在Nb感染期间前嗜碱性粒细胞在外周组织中可能出现的机制。
先前已经证明Nb感染能够引起全身性的IL-3上调,因此我们研究了IL-3对骨髓外出现的前嗜碱性粒细胞的可能贡献。在小鼠体内腹腔注射IL-3复合物(重组IL-3与单克隆抗IL-3抗体混合)后,外周血中出现CLEC12Ahi的前嗜碱性粒细胞(图. 6a)。我们推测IL-3诱导了前嗜碱性粒细胞从骨髓中外流。为了验证这一可能性,我们对IL-3和PBS对照处理的BMBAs进行了批量RNA测序分析,并鉴定了两者之间的差异表达基因(DEGs)。我们特别注意到Cxcr4基因的表达在IL-3刺激下显著降低(图. 6b),因为CXCR4已被证明是一种重要的趋化因子受体,对免疫细胞在骨髓中的滞留起作用。骨髓细胞的单细胞RNA测序和流式细胞分析显示,在转录和蛋白水平上,CXCR4在前嗜碱性粒细胞中的表达高于成熟嗜碱性粒细胞(图. 6c),而IL-3刺激能够降低CLEC12Alo成熟嗜碱性粒细胞和CLEC12Ahi前嗜碱性粒细胞表面CXCR4的表达(图. 6d)。这些结果表明,由于CXCR4的高表达,前嗜碱性粒细胞留在骨髓中,而IL-3介导的CXCR4下调可能促进前嗜碱性粒细胞从骨髓中外出。根据这一假设,给小鼠腹腔注射一种CXCR4抑制剂AMD3100,可导致外周血和脾中出现CLEC12Ahi前嗜碱性粒细胞(图. 6e)。因此,因此,在骨髓的前嗜碱性粒细胞中,IL-3介导的CXCR4的下调至少部分地解释了前嗜碱性粒细胞从骨髓中流出,并在Nb感染期间它们出现在肺和皮肤中。
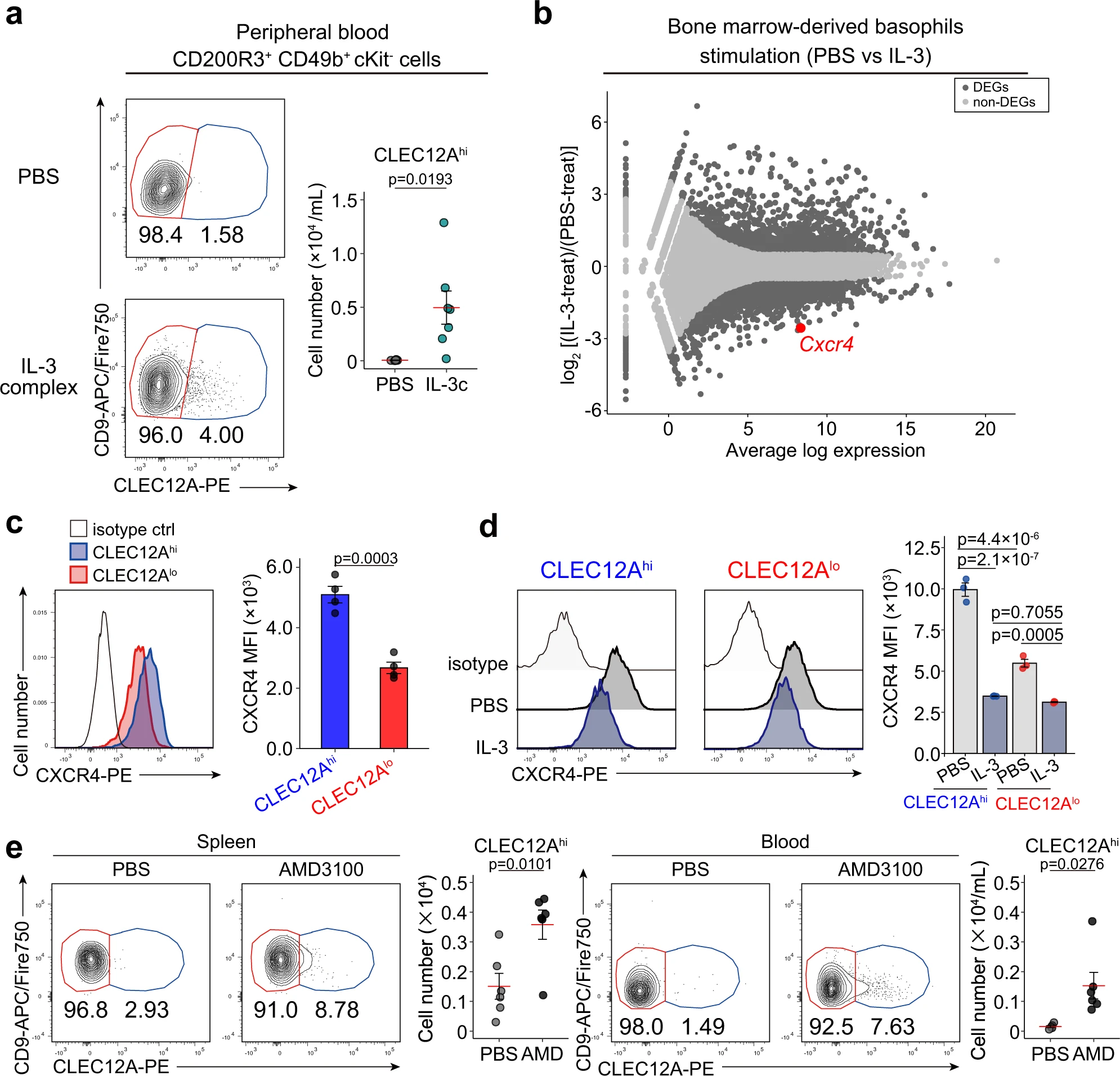
图6|前嗜碱性粒细胞中IL-3介导的CXCR4下调促进其从骨髓中排出
参考文献:
Miyake, K., Ito, J., Nakabayashi, J. et al. Single cell transcriptomics clarifies the basophil differentiation trajectory and identifies pre-嗜碱性粒细胞 upstream of mature 嗜碱性粒细胞. Nat Commun 14, 2694 (2023).https://doi.org/10.1038/s41467-023-38356-1