






单细胞和空间转录组分析揭示了肝转移性结直肠癌的细胞异质性
在这项研究中,作者使用单细胞和空间转录组RNA测序,全面描绘了结直肠癌(CRC)和配对良好的肝转移的细胞格局。作者从6例结直肠癌患者的27份标本中产生了41,892个CD45-非免疫细胞和196,473个CD45+免疫细胞,发现CD8_CXCL13和CD4_CXCL13亚群在肝转移样本中显著增加,表现出高增殖能力和肿瘤激活特性,有助于改善患者的预后。在原发瘤和肝转移瘤中观察到不同的成纤维细胞特征。原发肿瘤中富含F3+的成纤维细胞通过表达促肿瘤因子而导致总体生存率下降。而肝转移瘤中MCAM+成纤维细胞可能通过Notch信号途径促进CD8_CXCL13细胞的产生。综上所述,作者通过单细胞和空间转录组RNA测序,广泛分析了原发和肝转移结直肠癌细胞图谱的转录差异,提供了结直肠癌肝转移发生发展的不同维度。
该研究于2023年6月发表发表在《Science advances》,IF:14.957。
技术路线
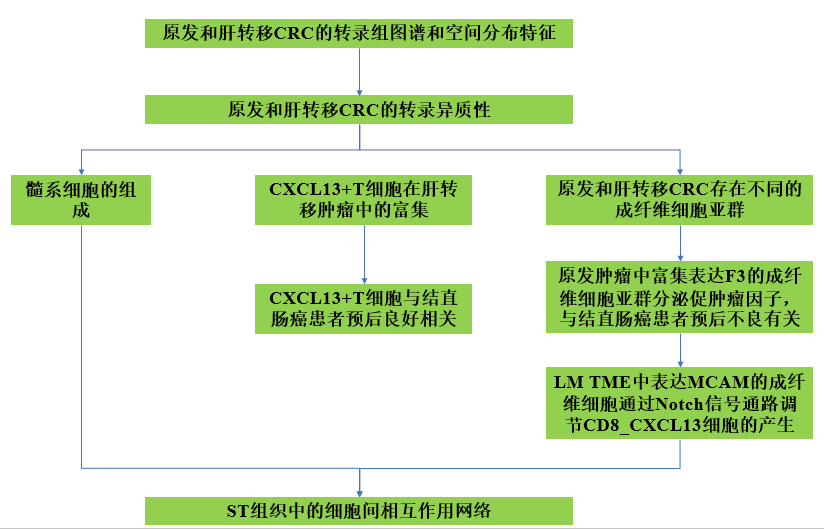
1、CRC原发肿瘤和CRC来源的肝转移肿瘤的总体单细胞转录组图谱
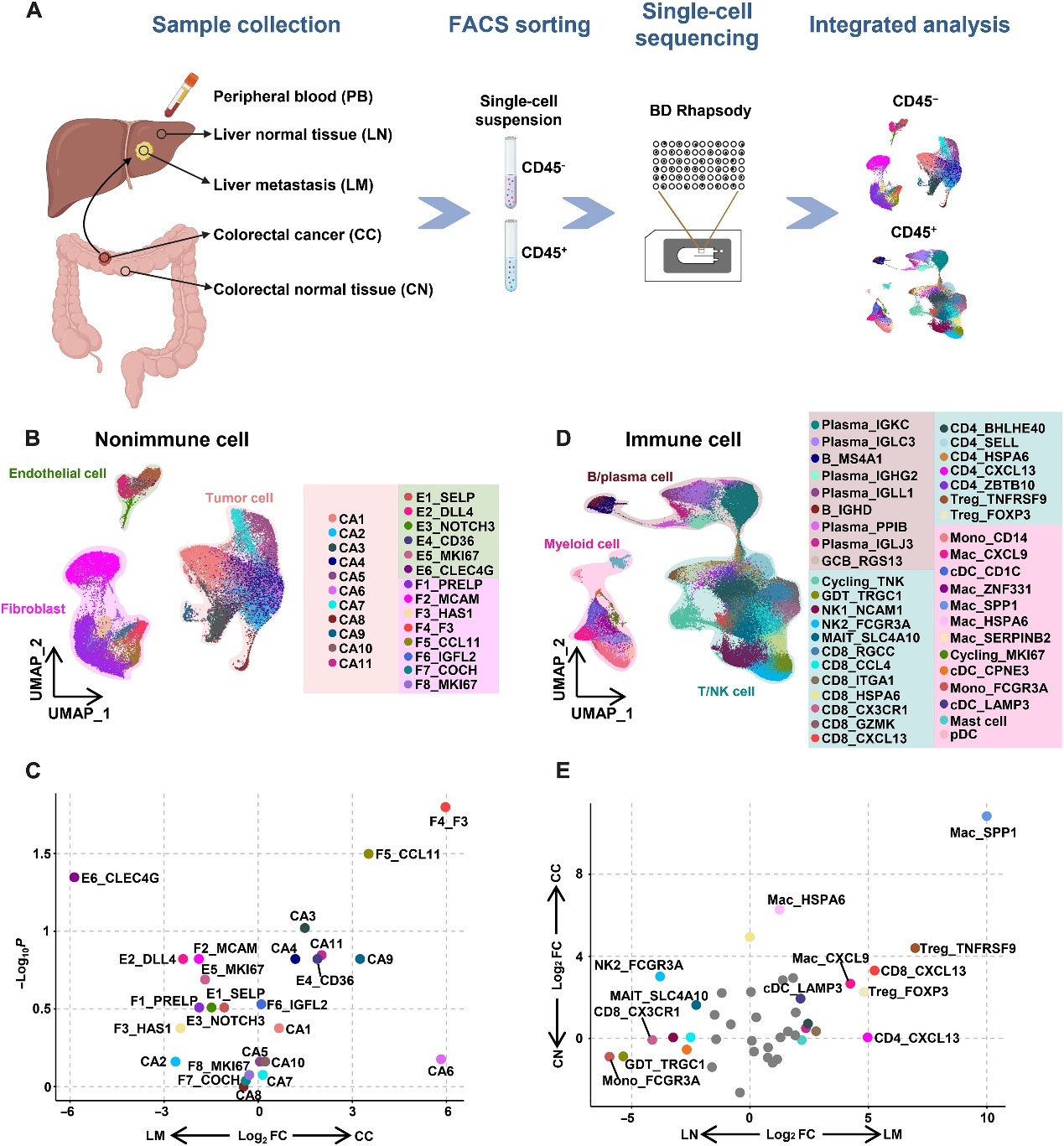
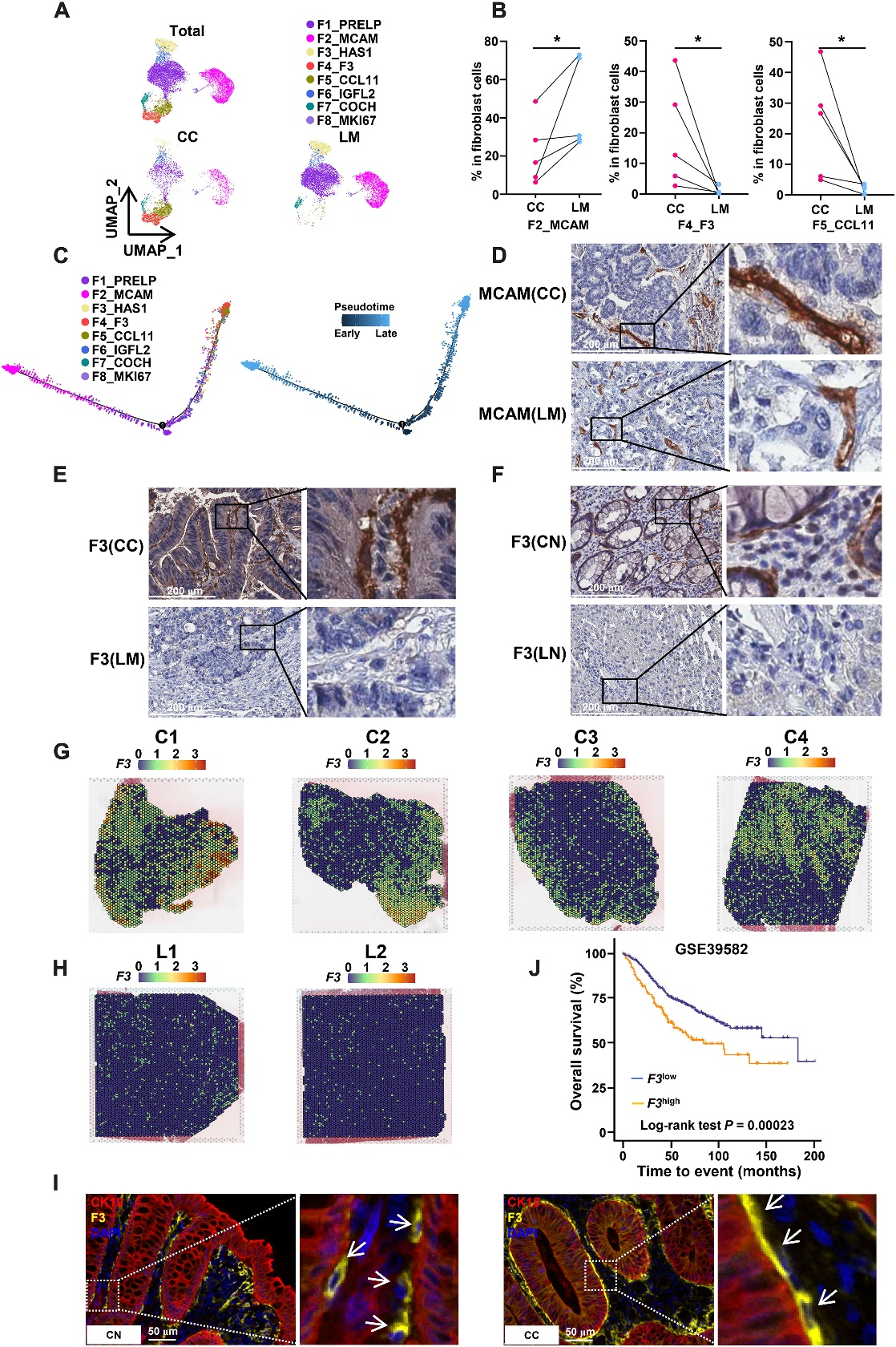
为了阐明CRC原发肿瘤和CRC来源的肝转移肿瘤的细胞异质性,作者从六名CRC患者的原发结直肠癌(CC)、相邻正常结直肠粘膜(CN)、肝转移(LM)、相邻正常肝组织(LN)以及外周血(PB)中收集了CD45−非免疫细胞和CD45+免疫细胞进行单细胞转录组分析(图1A)。经过质量控制和过滤,作者从27个样本中获得了41,892个CD45−非免疫细胞和196,473个CD45+免疫细胞,并鉴定出了23个非免疫细胞簇和41个免疫细胞簇。通过基于标记物的注释,作者从非免疫细胞中鉴定出了肿瘤细胞(EPCAM和SOX9)、成纤维细胞(COL1A1和COL1A2)和内皮细胞(PECAM1和CD34),而从免疫细胞中鉴定出了T细胞(CD3、pAbO)、自然杀伤细胞(NK细胞)(CD56、pAbO)、B细胞/浆细胞(CD19、pAbO)、单核细胞/巨噬细胞(CD14、pAbO)、树突状细胞(DCs)(HLA.DRA)和肥大细胞(TPSAB1)。
进一步对非免疫细胞主要群体进行无监督聚类分析,得到了11个肿瘤细胞簇、8个成纤维细胞簇和6个内皮细胞簇(图1B)。CA2和F2_MCAM在LM中富集。F4_F3和F5_CCL11在CC中显著富集(图1C)。基于已知标记物的表达情况,将免疫细胞分为41个亚群(图1D)。在原发肿瘤微环境(TME)中,调节性T细胞(Tregs)和巨噬细胞的百分比较CN对照组更高。虽然单核细胞、NK细胞和黏膜相关固有T细胞(MAIT细胞)在LN中占主导地位,但转移肿瘤细胞对Tregs和巨噬细胞的显著上调重塑了TIME(图1E)。总之,作者在CRCLM的不同组织中鉴定出多个具有不同分布模式的细胞亚群。
2、 CRC原发肿瘤和CRC来源的肝转移肿瘤的空间分布特征
为了全面分析CRC原发肿瘤和CRC来源的肝转移肿瘤的空间分布特征,作者从六名患者中收集了六个组织标本,包括四例CRC原发肿瘤(C1至C4)和两例肝转移肿瘤(L1和L2)。通过hematoxylin和伊红(H&E)染色以及每个样本的基因表达特征,识别出肿瘤区域(T)和肿瘤周围区域(PT)(图2A)。与肿瘤周围组织相比,肿瘤组织富集了与细胞周期相关的通路,如G2-M检查点、E2F靶点和有丝分裂纺锤体(图2B)。根据基因表达谱,将肿瘤组织和肿瘤周围组织分为不同的区域(图2A)。考虑到每个区域包含多个细胞,作者采用了一种基于标记物的策略,将空间转录组(ST)和单细胞RNA测序(scRNA-seq)数据整合起来,估计每个捕获区域中不同细胞类型的比例。在ST组织中鉴定出B细胞、T细胞、自然杀伤细胞(NK细胞)、浆细胞、髓系细胞、肿瘤细胞、成纤维细胞和内皮细胞。这些细胞簇的比例和得分在每个区域中有所变化。与C1、C3和C4的肿瘤周围组织相比,浆细胞的比例在肿瘤组织中下降(图2C和2D)。

3、原发和肝转移瘤肿瘤细胞转录异质性
研究表明,人类结直肠肿瘤细胞包含多种细胞类型,其转录特征反映了正常结直肠上皮的特征。在正常上皮中已经确定了干细胞样/转运增殖细胞、结肠细胞、腺细胞和毛刷细胞。进行了亚类聚类分析,并揭示了肿瘤细胞的不同分化谱系,即CA1到CA11。由单个肿瘤细胞产生的肿瘤组织可以概括出亲本肿瘤的细胞多样性。LM中的肿瘤细胞群体多样性与CC相似(图1C)。为了进一步研究不同肿瘤细胞簇的功能通路和转录程序,作者进行了基因集变异分析(GSVA)和转录因子(TF)分析。结果显示,CA2富集了Wnt-β-catenin信号通路,并在CA2中上调了在Wnt-β-catenin信号通路中起重要作用的转录因子LEF1。此外,CA2表现出体细胞干细胞分裂通路的上调。一致的是,CA2具有最高的LGR5表达水平,这是肠道干细胞的标记物。这些结果表明,CA2可能是TME中的干细胞亚群。此外,CA2具有EPCAM、CDH1和PRSS2的高表达水平,促进肿瘤细胞的粘附和定植。总的来说,聚类鉴定和特征分析展示了肿瘤细胞的功能多样化,转移肿瘤能够重现亲本肿瘤的细胞多样性。
4、TME重塑髓系细胞的组成
髓系细胞是TME中异质的亚群,作者鉴定出了三种髓系细胞:单核细胞/巨噬细胞、树突状细胞(DCs)和肥大细胞。通过亚类聚类分析,作者鉴定出了八个单核细胞/巨噬细胞亚群和三个传统DCs(cDCs)亚群(图3A),并且每个亚群都具有独特的基因表达模式。每个部位的髓系细胞具有不同的分布模式(图3B),展示了器官特异性的特征。CD14+单核细胞是PB中最富集的亚群。与肿瘤周围组织相比,Mac_SPP1亚群在肿瘤组织中富集,而大部分Mac_SPP1亚群细胞来自原发肿瘤。此外,Mac_CXCL9的比例在LM中显著增加(图3C),该亚群表达CXCL9的水平较高,表明该亚群能够招募CXCR3阳性效应T细胞进入肿瘤。Mac_SPP1亚群中富集参与招募髓系细胞,尤其是粒细胞如CXCL3的基因(图3D)。髓系抑制性细胞可以高表达CXCR2,CXCL3的受体,并且可以通过表达精氨酸酶和诱导型一氧化氮合酶(iNOS)来抑制CD8+T细胞。GSVA分析显示,Mac_SPP1亚群参与与炎症反应相关的途径。然而,Mac_CXCL9亚群在干扰素-γ(IFN-γ)反应和T细胞活化途径中富集(图3E)。轨迹分析显示,Mac_CXCL9和Mac_SPP1亚群都是终末分化的,表明它们是肿瘤活化的亚群(图3F)。
根据细胞因子表达和功能,DCs可以分为浆细胞样DCs(pDCs)和cDCs。在TME中已经鉴定出不同亚群的cDCs,包括cDC1和cDC2。在作者的数据中,CC和LM中都鉴定出了cDC1(cDC_CPNE3)和cDC2(cDC_CD1c)(图3A)。此外,在LM中还富集了cDC_LAMP3(图3C)。cDC_LAMP3表达CCR7的水平较高(图3G),CCR7是CCL19和CCL21的受体,表明它具有迁移至淋巴结的能力。此外,cDC_LAMP3亚群表现出CD40、CD80和CD86这些共刺激分子的高表达水平,这是DC成熟的标记物(图3H)。cDC_LAMP3亚群的百分比在LM中显著增加,表明TIME中肿瘤细胞的增长。

5、CXCL13+T细胞在肝转移肿瘤中的富集
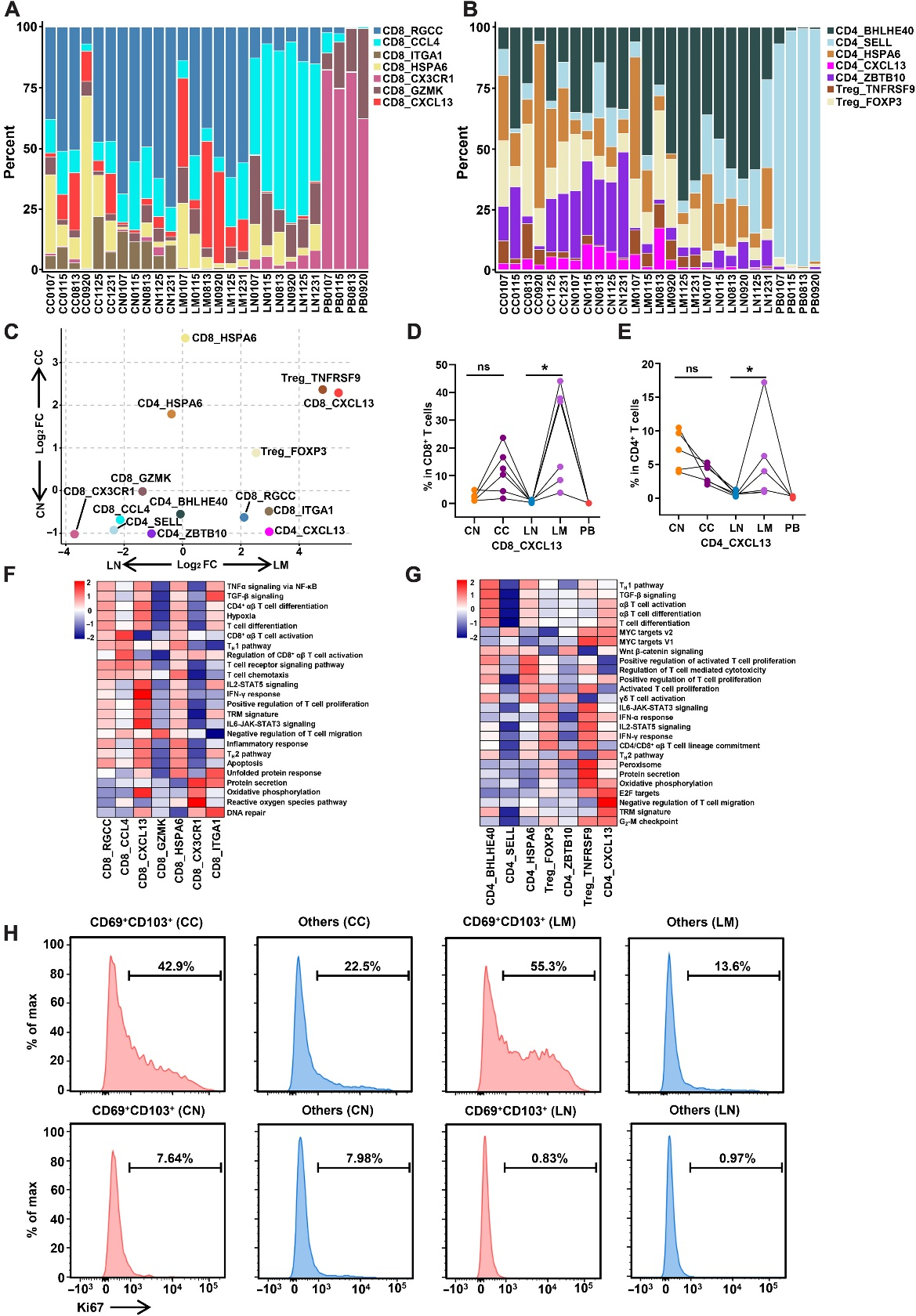
T细胞,特别是CD8+ T细胞和CD4+ T细胞,在适应性免疫应答中起主导作用。在T细胞中,鉴定出了七个CD8+ T细胞簇,五个传统CD4+ T(cCD4)细胞簇和两个调节性T细胞(Tregs)簇。CD8+ T细胞和CD4+ T细胞都包括表达CXCL13的簇,这是CXCR5的趋化因子(图4A、B)。与LN相比,CD8_CXCL13细胞的百分比在LM中显著增加;然而,在CN、LN和PB中很少检测到这个亚群(图4C和D)。在结肠的稳态下,大约3%至12%的CD4细胞为CXCL13阳性,这被称为滤泡性辅助T细胞(TFH)。在CC中,CD4_CXCL13的百分比下降,但与LN相比,在LM中这个亚群的百分比增加(图4E)。
为了进一步研究CXCL13+ T亚群的特征,作者分析了每个亚群中上调的通路。GSVA分析显示,CD8_CXCL13亚群富集了T细胞增殖通路,而CD4_CXCL13亚群富集了G2-M检查点通路,显示出这两个亚群的增殖特性(图4F和G)。每个簇的基因分析显示,CD8_CXCL13细胞表达ITGAE的水平较高,ITGAE是组织驻留记忆T(TRM)细胞的标记物。考虑到TRM细胞也是CD69阳性的,作者通过流式细胞术鉴定了CD69+CD103+CD8+ T细胞。大多数CD103+细胞是CD69阳性细胞,而CD69+CD103+CD8+ T细胞在LN中很少出现。一致地,来自CC和LM的CD69+CD103+CD8+ T细胞中Ki67的表达水平高于其他CD8+ T细胞。来自CN和LN的不同CD8+ T细胞亚群的增殖特性几乎相同(图4H)。综上所述,这些结果表明,由于其高增殖能力,CD8_CXCL13和CD4_CXCL13细胞在CRC的LM中上调。
6、CXCL13+T细胞与结直肠癌患者预后良好相关
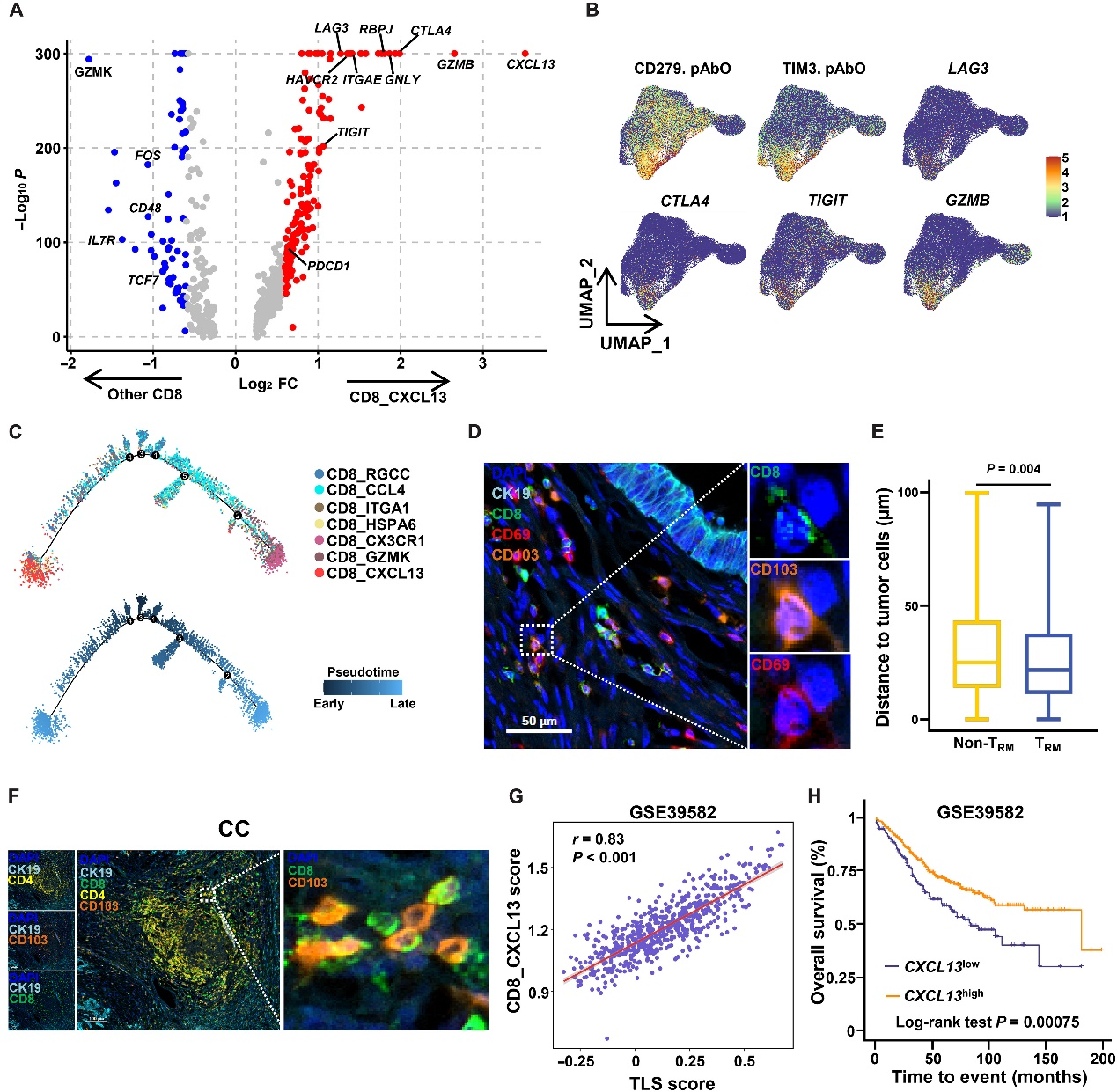
在上述结果中,作者展示了与CD4_CXCL13亚群相比,CD8_CXCL13在CC和LM中富集(图4D和E),这表明CD8_CXCL13细胞可能是一种激活肿瘤的亚群。因此,作者进一步研究了CD8_CXCL13亚群的特征。与其他亚群相比,CD8_CXCL13亚群中观察到了PDCD1、HAVCR2、LAG3、CTLA4和TIGIT等耗竭标记物的高水平(图5A和B)。先前的研究显示,由于持续的肿瘤抗原刺激,肿瘤反应性T细胞表现出衰竭表型,而衰竭表型表明它们的肿瘤反应性。轨迹分析显示,CD8_CXCL13细胞是终末分化的(图5C)。此外,CXCL13+ T细胞亚群在TME中表现出显著的克隆扩增能力,进一步表明它们具有抗原经历的特性。此外,CD8_CXCL13亚群还表达高水平的效应分子,如GZMB(图5A和B),这表明该亚群可能保留了部分抗肿瘤功能。由于在组织中检测CXCL13较为困难,作者尝试使用CD69和CD103来标记CXCL13+细胞,免疫组织化学(IHC)结果显示,在肿瘤组织中,CD69+CD103+CD8+ T细胞与CK19+肿瘤细胞更加接近(图5D和E),有利于它们的抗肿瘤功能。值得注意的是,CD103+CD8+ T细胞存在于结直肠癌组织的第三淋巴结结构(TLS),TLS评分较高的患者CD8_CXCL13评分也较高,这表明该亚群可能参与了TLS的形成(图5F和G)。
为了探索CXCL13+ T细胞在结直肠癌中的预后价值,作者将来自基因表达库(GEO)队列的结直肠癌患者分为CXCL13高表达组和CXCL13低表达组,并发现CC中CXCL13的高表达预示着更好的总体生存率(图5H)。总之,富集在TME中的CXCL13+ T细胞是一种对肿瘤具有反应性的亚群,并有助于结直肠癌患者的良好预后。
7、原发和肝转移结的直肠癌中存在不同的成纤维细胞亚群
成纤维细胞是TME中非免疫细胞的主要类型。作者进一步表征成纤维细胞,探索它们在原发结直肠癌和肝转移肿瘤中的异质性。在结直肠癌的TME中鉴定出八个具有独特基因表达模式的成纤维细胞亚群,分别被命名为F1_PRELP、F2_MCAM、F3_HAS1、F4_F3、F5_CCL11、F6_IGFL2、F7_COCH和F8_MKI67(图6A)。F2_MCAM亚群在LM中的比例高于CC。F4_F3和F5_CCL11亚群大部分由CC来源的细胞组成(图6B)。根据批量RNA测序数据,F4_F3的浸润在CC中增加,与CN相比。然而,根据轨迹分析,F5_CCL11亚群在CC中的比例降低,可能转变为F4_F3亚群(图6C)。轨迹分析还预测F2_MCAM和F4_F3是两个不同的终末分化亚群(图6C)。IHC分析验证了F2_MCAM在CC和LM中的存在(图6D)。根据IHC结果,MCAM+CAF在LN中非常罕见。大部分MCAM+细胞是内皮细胞。然而,与单细胞分析一致,F4_F3亚群只存在于CC中,而在LM中不存在,这可能是由于LN中缺乏F4_F3而CN中存在的现象(图6E和F)。ST分析也证实,在CC中F4_F3亚群的浸润要比LM中多(图6G和H)。F4_F3亚群紧密包围CN和CC中的上皮细胞,促进其与上皮细胞的相互作用(图6I)。此外,F4_F3在CC中的增加可能导致更差的预后(图6J)。总之,作者观察到原发结直肠癌和肝转移肿瘤之间的成纤维细胞具有不同的表型特征和高度可变的频率,这表明在不同癌症环境中TME内存在细胞异质性。
8、原发肿瘤中富集表达F3的成纤维细胞亚群分泌促肿瘤因子,与结直肠癌患者预后不良有关
为了研究在CC和LM中富集的不同成纤维细胞对TME的重塑作用,作者进一步分析了F2_MCAM和F4_F3的特征。F2_MCAM富集了JAG1和NOTCH3,这两者参与了NOTCH信号通路。F4_F3富集了C3和CXCL1,表明该亚群参与了补体和炎症反应通路(图7A和B)。F4_F3还高表达MMP2和MMP3,这可能与细胞外基质的组织结构有关(图7A)。此外,F4_F3还富集了参与血管生成和肿瘤浸润的促肿瘤因子,如VEGFA、NRG1、HGF、GDF15、AREG和BMP2(图7C)。F4_F3与肿瘤细胞之间的相互作用分析进一步揭示了它们通过NRG1和Erb-B2受体酪氨酸激酶3(ERBB3)信号通路进行交流,而ERBB3几乎富集在肿瘤细胞中,并可以与ERBB2形成异源二聚体,促进肿瘤细胞增殖并使CRC患者对西妥昔单抗产生耐药性(图7D)。ST分析还显示了F3与NRG1的共定位,围绕着ERBB3+肿瘤细胞(图6G和图7E至I)。跨膜迁移实验显示重组人NRG1(rNRG1)可以促进RKO和SW620细胞的迁移(图7J)。高表达F3和NRG1的CRC患者预后较差(图7K)。这些结果表明,富集于CC的F4_F3成纤维细胞通过分泌促肿瘤因子可能导致CRC患者预后不佳。

9、LM TME中表达MCAM的成纤维细胞通过Notch信号通路调节CD8_CXCL13细胞的产生
在CD8_CXCL13和CD4_CXCL13亚群中,Notch信号通路的转录因子RBPJ表达较高(图5A)。有报道称,Notch信号通路可以通过配体和受体的相互作用来激活。在与其配体相互作用后,Notch的胞内结构域可以被切割并转位到细胞核中,调控其靶基因的转录。LM中RBPJ的表达与CXCL13和ITGAE呈正相关,而在CC中未观察到这种关联(图8A)。CD8_CXCL13和CD4_CXCL13亚群主要表达NOTCH1受体(图8B)。为了进一步了解哪些细胞类型在CD8_CXCL13和CD4_CXCL13亚群中调控Notch信号通路,作者分析了Notch配体的表达水平,包括DLL1、DLL3、DLL4、JAG1和JAG2,发现Notch配体主要在成纤维细胞和内皮细胞中表达。F2_MCAM亚群富集了JAG1,F5_COCH亚群富集了DLL1,而E2_DLL4亚群富集了DLL4、JAG1和JAG2(图8C)。使用CellPhone DB进行的Notch及其配体的相互作用分析显示,在内皮细胞中,E2_DLL4亚群与CXCL13+ T细胞之间的相互作用最强,而在成纤维细胞中,F2_MCAM亚群通过JAG1-NOTCH1与CD8_CXCL13和CD4_CXCL13亚群相互作用(图8D)。由于成纤维细胞在TME中的分布呈散点状,作者推测F2_MCAM亚群有助于激活CXCL13+ T细胞中的Notch信号通路。
接下来,作者根据LM中F2_MCAM的比例将患者分为两组,并发现F2_MCAM高的LM中CD8_CXCL13亚群比例较高(图8E)。通过GEO数据集分析发现,F2_MCAM浸润评分较高的患者在LM中CD8_CXCL13亚群的浸润评分也较高,但在CD4_CXCL13亚群和CC中未观察到类似的关联(图8F)。此外,作者通过ST检测了LM中F2_MCAM和CD8_CXCL13的位置。与单细胞结果一致,F2_MCAM浸润评分较高的区域在ST样本中CD8_CXCL13的浸润评分也较高(图8G和图8H)。此外,LM中Notch信号通路的相互作用强度比CC中更强,这可能是由于LM中F2_MCAM亚群的比例较高(图6B)。
已有研究报道Notch信号通路调控CD8+ T细胞的抗肿瘤免疫,但对于Notch信号通路是否可以调控CXCL13的表达了解甚少。为了探索这种调控作用,作者使用JASPAR(http://jaspar.genereg.net/)预测了RBPJ在CXCL13启动子上的结合位点。发现了几个潜在的结合位点,表明RBPJ可能作为转录因子影响CXCL13的表达。

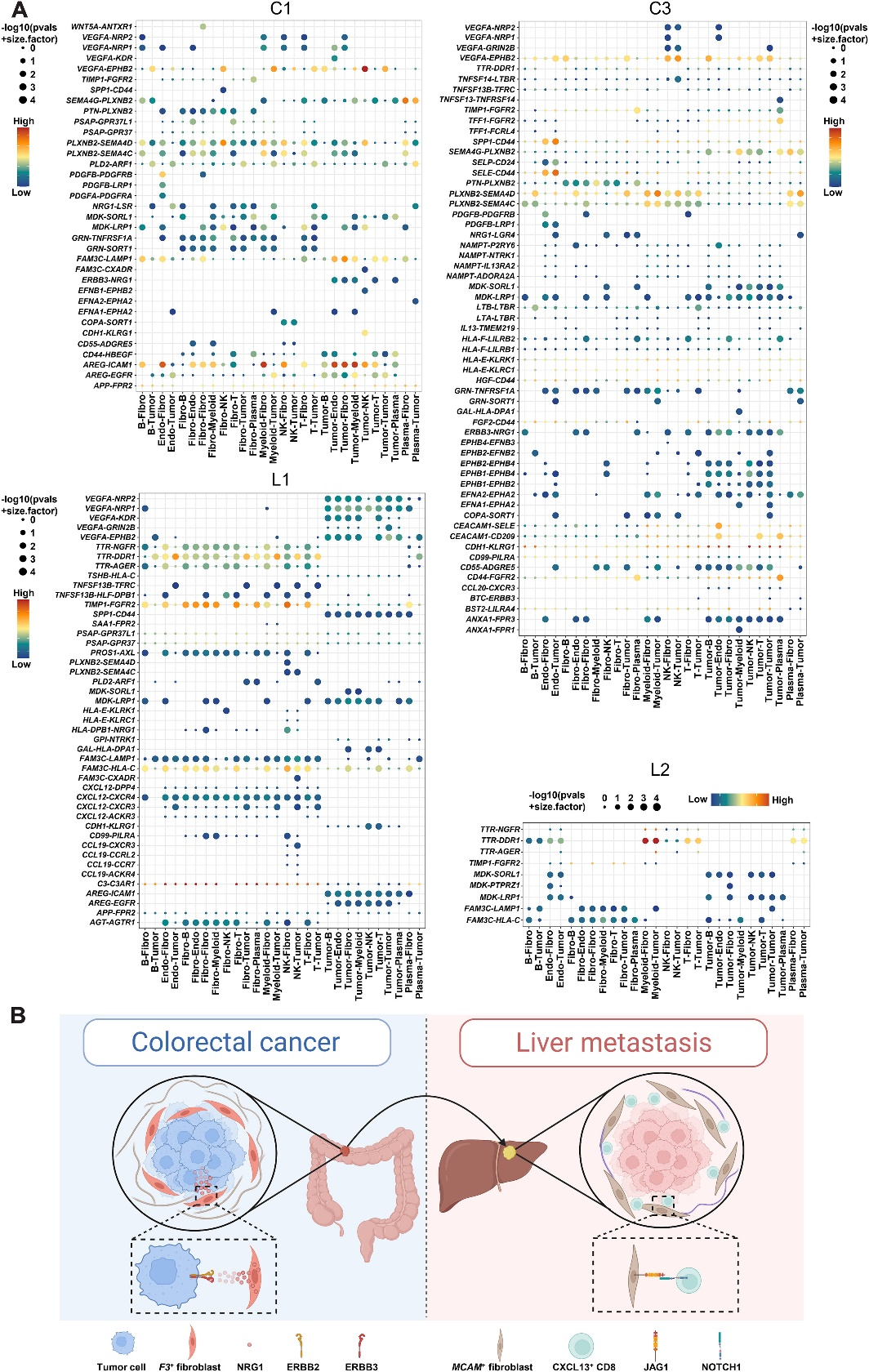
10、ST组织中的细胞间相互作用网络
作者还研究了原发肿瘤和肝转移瘤的ST中不同簇之间的细胞相互作用。结果显示,VEGFA-NRP1和VEGFA-NRP2配体受体对在原发肿瘤和肝转移瘤中都富集。作者还发现CC和LM之间存在不同的富集配体受体对。ERBB3-NRG1配对在C1和C3中富集,但在L1和L2中不存在(图9A),这表明ERBB3-NRG1相互作用在原发肿瘤的发展和转移中可能具有潜在作用。
综上所述,作者发现在CRC原发肿瘤中富集的F3+成纤维细胞可以通过产生包括NRG1在内的各种促肿瘤因子来调节肿瘤的发展和/或迁移,这些因子与肿瘤细胞表达的ERBB3相互作用以发挥其促肿瘤功能。然而,在LM中富集的MCAM+成纤维细胞可以通过Notch信号通路调节CD8_CXCL13细胞的生成,与CC中的促肿瘤亚群F3+成纤维细胞不同,这表明不同癌症环境中间质细胞的细胞异质性(图9B)。
实验方法
组织加工与细胞分选、scRNA-seq、无监督的细胞聚类和标注、ssGSEA、GSVA、SCENIC、Monocle2、Spatial-seq、流式细胞术、CellPhone DB、IHC染色、生存分析、基因表达相关性分析、Transwell迁移率分析。
参考文献
Wang F, Long J, Li L, Wu ZX, Da TT, Wang XQ, Huang C, Jiang YH, Yao XQ, Ma HQ, Lian ZX, Zhao ZB, Cao J. Single-cell and spatial transcriptome analysis reveals the cellular heterogeneity of liver metastatic colorectal cancer. Sci Adv. 2023 Jun 16;9(24):eadf5464. doi: 10.1126/sciadv.adf5464. Epub 2023 Jun 16.