






帕金森病大脑的转录组学分析揭示疾病阶段特异性基因表达变化

帕金森病(PD)是常见的仅次于阿尔茨海默病的神经退行性疾病,65岁以上人群中有2-3%的人患病。目前还没有有效减缓疾病进程的治疗方法,迫切需要更好地了解疾病机制以支持新疗法的开发。PD的特征是黑质致密部多巴胺能神经元的神经变性,导致纹状体多巴胺耗竭,引起静止性震颤、运动迟缓和强直等症状。另一个神经病理学标志是在剩余神经元和神经元突起中分别存在路易小体(LB)和路易神经突包涵体,主要由错误折叠的α-突触核蛋白(α-syn)聚集物组成。2003年Braak及其同事基于脑内LB病理的特定进展模式提出了临床前和临床PD病理分期系统。Braak LB分期范围从1到6,反映出大脑中LB的分布越来越广泛,呈现出从头端到尾端的模式。Braak分期系统被认为反映了疾病进展,随着疾病过程进一步影响脑区,关键临床症状相继出现。大多数PD是散发性的,可能是由遗传、表观遗传和环境之间的复杂相互作用引起的。全基因组关联研究发现了90个与PD风险增加相关的基因位点。表观遗传学和转录组学研究主要在血液和死后脑组织中进行,后者可以说是该疾病核心病理学最具代表性。然而,以往的PD脑转录组学研究在样本数量和所选脑区方面都受到限制。没有研究使用神经病理学,特别是Braak LB分期作为疾病进展的标志物。该研究发表在《Acta Neuropathologica》,IF:12.7。
技术路线

主要研究结果
1. 不同病理疾病阶段个体额上回的基因表达
在该研究中,作者检测了84个个体(包括23个非神经系统对照和61个在不同Braak LB分期受α-syn相关病理学影响的个体)的上额叶皮质转录组水平的差异。后者根据Braak LB分期分为三个神经病理组:一组由分析组织中没有LB病理的个体组成,对应Braak LB 1-4期,(n=19),一组由额叶皮层中度LB病理学的个体组成(Braak LB 5期,n=19),另一组由额叶皮层LB负荷较高的个体组成(Braak LB 6期,n=23)。值得注意的是,作者从额叶上皮层提取了RNA,LB在Braak LB 5期出现(图1)。
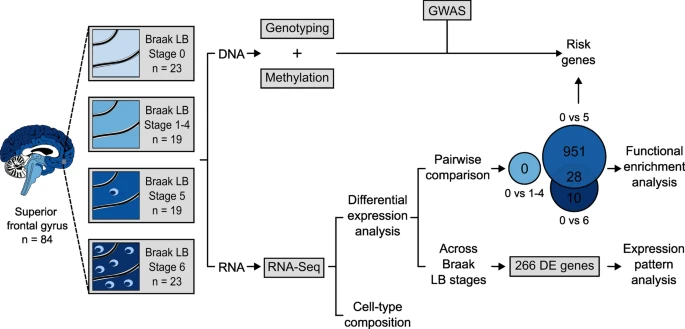
图1 研究概述
所选个体在人口统计学因素(性别和死亡年龄)和阿尔茨海默病(AD)相关病理水平相匹配(图2)。观察到接受深部脑刺激手术的个体比例在两组间有显著差异(Braak LB 0=0/23,Braak LB 1-4=0/19,Braak LB 5=1/19,Braak LB 6=6/23,p=0.0036,卡方检验)。这种差异可以用以下事实来解释:深部脑刺激可能是较晚期特定患者的一种治疗方案。组间痴呆持续时间也有差异,第5组和第6组的痴呆持续时间较长(p=0.0002,Kruskal-Wallis秩和检验)。这种差异可归因于痴呆出现在疾病的晚期阶段。值得注意的是,在Braak神经原纤维缠结期、反映总AD相关病理学的复合“ABC评分”、CERAD分期或CAA类型方面,两组间未发现差异,这表明伴随的AD病理不是作者分析中的重要混杂因素。此外,所有样本的死后延迟非常短(平均=377分钟,标准差=135分钟),确保了高质量的样本。
作者在去除核糖体RNA后进行了RNA测序,在所有样本中共检测到21,615个表达的基因,包括长链非编码RNA(lncRNA)。

图2 个体的人口统计学、样本信息和病理测量
2. 细胞组成和RNA质量
在作者的研究中,使用Scaden估计了细胞类型的比例。作者检验了细胞类型比例与数据中生物学变异的潜在来源(神经病理分组、性别、死亡时年龄、PMD和RIN)之间的Pearson相关性。神经元的比例与其他细胞类型的比例呈显著的抗相关(p<0.05)(图3a)。此外,如既往研究所示,细胞类型比例与RIN值呈正相关(与神经元比例呈正相关,与其他细胞类型比例呈负相关)(图3a)。细胞类型比例和其他变量(神经病理组、性别、年龄和PMD)之间没有发现关联(图3a)。然后,作者对已知的实验协变量(性别、死亡年龄、PMD、RIN)进行校正,寻找神经病理学组之间细胞类型比例的差异。在4个神经病理组(0、1-4、5和6)中,任何细胞类型均无显著差异(p<0.2)(图3b)。为了证实这一结果,作者也使用标记基因谱估计了样本中的细胞组成,发现神经病理组(0、1-4、5和6)之间任何细胞类型都没有显著差异(p<0.3)。
由于RNA质量与细胞类型比例相关,而且它也被证明影响基因表达水平测定,因此作者使用质量替代变量分析(qSVA)框架来估计RNA质量混杂因素。采用该方法获得的5个质量替代变量(qSVs)与已知的与RNA质量相关的变量(RIN和PMD)进行Pearson相关性分析。此外,由于作者已经表明细胞比例与由RIN值测量的RNA质量显著相关,作者还检验了5个qSV和细胞类型比例之间的Pearson相关性。qSV1与所分析的所有变量显著相关。除PMD与qSV2和qSV3显著相关外,qSV5与其他变量均显著相关。最后,qSV4与RIN值显著相关(图3c)。因此,作者纳入了5个质量替代变量作为协变量,而不是使用样本的RIN值和PMD。
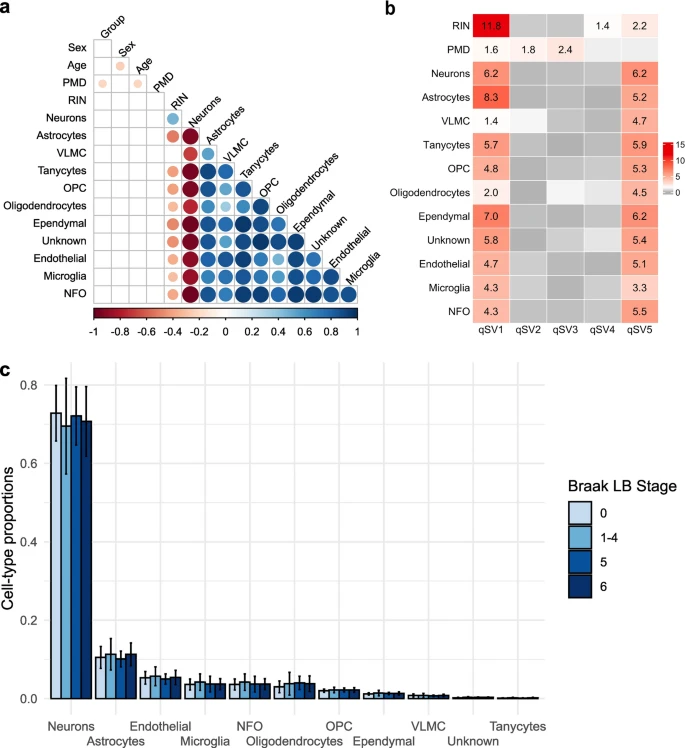
图3 细胞组成和RNA质量
3. 不同神经病理分期的差异基因表达
为了确定表达水平随LB神经病理学进展而变化的基因组,作者对所有4个神经病理学组的共21,615个转录本进行了差异基因表达分析。性别、死亡年龄和5个qSV被用作LRT检验的实验协变量。原假设是基因表达在组间是稳定的,并且检验与组顺序无关,这可能会检测出任何可能的上调和下调组合的不稳定表达模式。差异基因表达分析发现四个神经病理组共有266个差异表达基因(LRT检验,Benjamini-Hochberg FDR<0.05),其中34个对应于lncRNA。
为了进一步研究这266个差异表达基因,使用DEGreport R包中的分裂层次聚类对其正则化log2转换计数进行聚类。差异表达基因被分为8个簇。每个簇的典型神经病理组的表达模式,以及每个基因的缩放表达水平见图4。

图4 不同Braak LB分期的差异表达基因簇
簇1(30个基因)和4(25个基因)的表达分别从Braak LB 0期到6期呈线性上升和下降模式。上调基因簇(1)的命中率最高的基因是SNX7,而下调基因簇(4)的命中率最高的基因是NUCB1。所有其他的基因簇表现出一种表达模式,在Braak LB 1-4期和6期的样本有相对相似的表达水平,而在Braak LB 5期的样本与其他组相比显示出基因表达水平的重大变化。在这些显示主要转录组变化的簇中,簇2和3包含了大部分基因(分别为57和109个)。簇2中最重要的基因是PDXK,而簇3中最重要的三个基因是新的转录本。
4. Braak LB 0期对照和Braak LB病理组之间的两两差异基因表达
为了进一步研究Braak LB 5期样本的主要基因表达变化,作者比较了该组的转录组和对照组(Braak LB 0期)的转录组。使用Wald检验对共21,615个基因进行了差异基因表达分析,使用性别、死亡年龄和5个qSV作为协变量。作者发现在Braak LB 0期和Braak LB 5期样本之间共有979个差异表达基因(Benjamini-Hochberg FDR<0.05),其中575个表达上调,404个表达下调。在Braak LB 5期差异表达的979个基因中,233个基因在主要分析中比较了所有Braak LB分期组的基因表达差异的266个基因中。值得注意的是,在比较0期和1-4期时,没有发现在FDR<0.05的差异表达基因,而在Braak LB 0期和6期之间有38个基因有显著差异表达。在这38个在Braak LB 0期和6期之间有差异表达的基因中,有28个也在Braak LB 0期和5期之间的差异表达基因中。在Braak LB 5期和6期均以SNX7基因为主。这些发现进一步强调了在Braak LB 5期可以观察到最显著的额叶皮质基因表达变化。
5. 功能富集分析
作者对每个神经病理组(1-4、5和6)与对照组的差异基因表达结果进行了功能富集分析。作者纳入了所有21,615个基因,并根据每次比较的p值和对数倍数变化对每个基因进行评分。通过这种方法,作者发现从Braak LB 0期到1-4期显著富集了99条GO通路(FDR<0.05),从Braak LB 0到5期显著富集了75条通路,从Braak LB 0到6期显著富集了163条通路。作者生成了火山图,突出了在每个Braak LB分期富集的三个最重要的通路所涉及的基因。所有通路的调控方向是由每个通路所涉及的大多数基因驱动的。这排除了通路发现是由单基因表达变化驱动的可能性。在总结GO通路时,作者发现在Braak LB 1-4期富集的通路有29条,在Braak LB 5期富集的通路有31条(图5),在Braak LB 6期富集的通路有39条。此外,作者发现在所有Braak LB分期富集的通路有12条(均为下调),这些通路主要涉及ATP代谢过程。
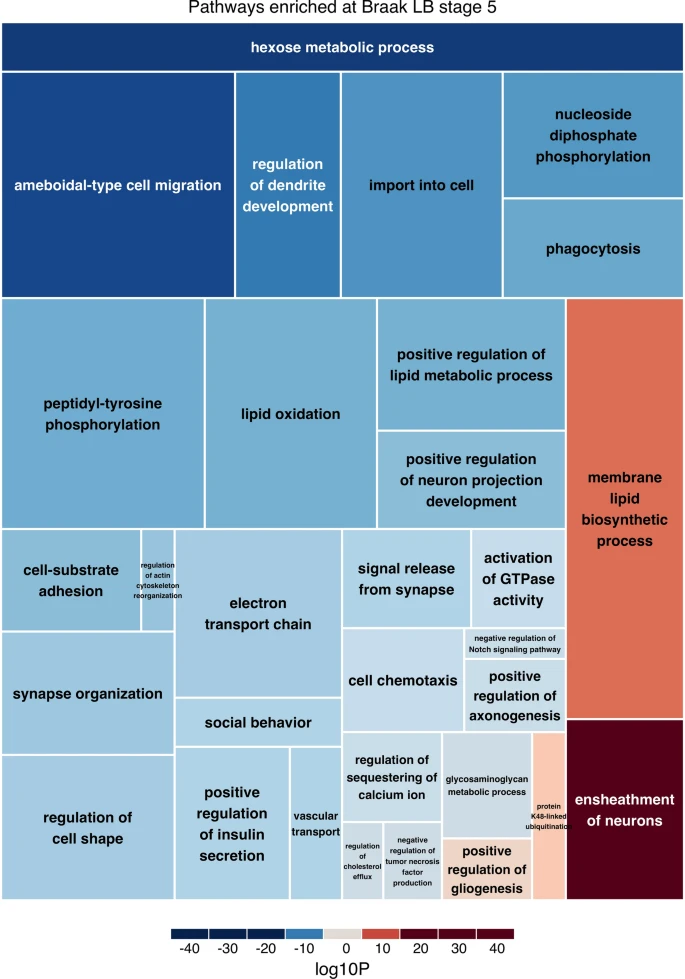
图5在Braak LB 5期富集的通路
6. 风险基因
为了研究部分SNPs与PD相关的功能和分子机制,作者提取了与90个帕金森病风险SNPs最近的基因和QTL指定基因。选取的98个基因中有87个在样本中表达。在Braak LB 0期和Braak LB 5期的差异表达基因中有5个(Benjamini-Hochberg FDR<0.05)。在Braak LB 5期的差异表达基因中,PD基因的富集没有统计学意义(X2(1,n=21,615)=0.084,p=0.773)。为了进一步研究这些基因,作者进行了eQTL分析,以评估这5个基因的表达是否与附近PD风险SNP的基因型相关。作者发现rs11683001与其最近的基因MAP4K4之间存在关联(校正后p=0.025)(图6a)。值得注意的是,在样本中表达的87个风险基因都不在Braak LB 0期和Braak LB 6期之间差异表达的基因中。
作者最近报道了与TMCC2、SFMBT2、AKAP6和PHYHIP附近的与Braak LB分期相关的差异甲基化复制位点。作者发现所有4个基因都在样本中表达。其中一个位于cg04011470附近的基因PHYHIP也是Braak LB 5期的差异表达基因之一(图6b)。

图6 风险基因
结论
综上所述,作者对Braak LB各阶段的转录组学分析表明,只有当LB首次出现在研究的病变组织中时,才能观察到基因表达的主要变化,而在LB出现之前和之后,只有微小的变化可以检测到。此外,作者证实了参与ATP代谢过程和突触损伤的通路在疾病的各个阶段都有富集。相反,与免疫反应相关的通路在LB形成之前被上调,然后在疾病的后期被下调,突显了免疫反应是未来疾病修饰治疗的可能靶点之一。作者还发现了可能参与PD发病的单个基因(SNX7,NUCB1,PDXK,PHYHIP和MAP4K4),并指出了lncRNA在疾病发病机制中的潜在作用。
实验方法
RNA测序
参考文献
Cappelletti C, Henriksen SP, Geut H, Rozemuller AJM, van de Berg WDJ, Pihlstrøm L, Toft M. Transcriptomic profiling of Parkinson's disease brains reveals disease stage specific gene expression changes. Acta Neuropathol. 2023 Jun 22. doi: 10.1007/s00401-023-02597-7.