






靶向m6A阅读器YTHDF1增强结直肠癌的抗肿瘤免疫和抗PD-1疗效
摘要
N6-甲基腺苷(m6A)在肿瘤免疫微环境(TIME)中的作用仍有待进一步研究。在这里,作者阐明YTHN6-甲基腺苷RNA结合蛋白1(YTHDF1)在大肠癌(CRC)TIME中的功能和机制。在组织微阵列(N=408)和TCGA(N=526)队列中评估YTHDF1的临床意义。在同基因肿瘤、肠特异性Ythdf1敲入小鼠和人源化小鼠中测定YTHDF1功能。采用单细胞RNA-seq(scRNA-seq)分析TIME。采用甲基化RNA免疫沉淀法测序(MeRIP-seq)、RNA测序(RNA-seq)和核糖体测序(Ribo-seq)鉴定YTHDF1直接靶标。用囊泡状纳米颗粒(VNPs)包封YTHDF1-siRNA进行YTHDF1体内沉默。在TCGA-CRC中,YTHDF1表达与干扰素-γ基因特征负相关。与此同时,在独立的组织微阵列队列中,YTHDF1蛋白与CD8+ T细胞浸润呈负相关,暗示其在TIME中的作用。基因缺失Ythdf1增强CT26(MSS-CRC)和MC38(MSI-HCRC)同基因肿瘤的抗肿瘤免疫,而在偶氮甲烷-葡聚糖-硫酸钠或ApcMin/+模型中,Ythdf1敲入蛋白促进免疫抑制TIME促进结直肠癌。scRNA-seq发现,在Ythdf1基因敲除肿瘤中,髓源性抑制细胞(MDSCs)减少,同时细胞毒性T细胞增加。综合MeRIP-seq、RNA-seq和核糖核酸-seq发现p65/Rela是YTHDF1的靶标。YTHDF1促进p65翻译上调CXCL1,从而增加MDSC通过CXCL1-CXCR2轴的迁移。增加的MSDCs反过来在时间上拮抗功能性CD8+ T细胞。重要的是,通过CRISPR(Clustered Regularly Interspaced Short Palindromic Repeats)或VNPs-siYTHDF1靶向YTHDF1可提高MSI-HCRC的抗PD1疗效,并克服MSSCRC的抗PD1耐药性。YTHDF1通过M6A-P65-CXCL1/CXCR2轴损害抗肿瘤免疫,促进结直肠癌,并在免疫检查点阻断治疗中作为治疗靶点。本文于2023年8月发表在《Gut》IF:24.5期刊上。
技术路线

主要实验结果
1、YTHDF1与结直肠癌中IFN-γ相关基因特征降低和预后不良相关
通过使用TCGA数据分析21个已知的m6A调控子的拷贝数变化,作者鉴定出YTHDF1是m6A调控子在近80%的CRC肿瘤中显示拷贝数增加或扩增中最高。这种拷贝数的增加也与mRNA表达和蛋白表达的上调相一致,提示YTHDF1具有促进CRC的功能。为建立抗肿瘤免疫与m6A调节因子之间的联系,该研究随后使用GSEA分析m6A调节因子与IFN-γ反应基因标记之间的相关性。作者发现YTHDF1与IFN-γ反应通路呈显著负相关(q<0.0001)。值得注意的是,IFN-γ反应与抗肿瘤免疫的诱导和对ICB治疗的反应有关。一致地,YTHDF1的表达与18 IFN-γ相关的基因标记呈强反相关(图1A),这预测多种癌症类型的抗PD1反应性。此外,CD8A或CD8+ T细胞特征的表达与YTHDF1表达呈负相关。与TCGA数据一致,来自CRC TMA队列的TMA IHC染色显示,队列I(p<0.001,r= -0.248,n=206)(图1B)和队列II(p<0.0001,r= -0.269,n=202)中,YTHDF1蛋白的高表达与CD8+ T细胞的低浸润相关(图1C)。这些数据强烈暗示,m6A阅读器YTHDF1与抗肿瘤免疫功能受损和ICB治疗效果降低有关。
与CD8+ T细胞浸润低与预后不良相关的观点一致,YTHDF1高表达(54.3%,100/184)预测CRC患者生存不良(p<0.01,log-rank检验)。多因素Cox回归分析证实,YTHDF1是队列II中结直肠癌的独立预后因素(HR,1.764;95%可信区间1.058-2.939;p<0.05)。在队列I中,通过多变量Cox回归分析进一步验证这些发现。
2、单细胞转录组学揭示YTHDF1诱导的免疫抑制
为研究YTHDF1在调节抗肿瘤免疫中的作用,作者使用CRISPR-Cas9系统敲除MC38小鼠MSI-H CRC细胞中的Ythdf1(Ythdf1-KO),并将细胞注射到同基因C57BL6小鼠(图1D)。发现与对照组(NC)相比,敲除Ythdf1后,肿瘤体积和重量都减少(图1E)。为了解Ythdf1-KO是否影响TIME,作者从肿瘤中分离出CD45+免疫细胞,并进行单细胞RNA-seq(scRNA-seq)(NC:1480个细胞;Ythdf1-KO:1816个细胞)。与NC组相比,含有Ythdf1-KO的肿瘤表现出粒细胞髓源性抑制细胞(G-MDSCs,簇3)和中性粒细胞的显著减少(图1F)。相比之下,Ythdf1-KO肿瘤中T细胞和NK细胞大量增加(图1F)。作者进一步将T细胞和NK细胞重新聚集为CD4+ T、CD8+ T、NKT和NK细胞亚群,并发现与对照组相比,它们在Ythdf1-KO肿瘤中同时增加(图1F)。因此,作者推测YTHDF1可以通过诱导MDSC积累来抑制抗肿瘤免疫。通过检测MDSCs的功能标记物Il1b、Arg2、Cxcr2和Ccr2,这些基因主要富集于MDSC簇(簇3和簇4),特别是来自未敲除Ythdf1的肿瘤(图1G)。因此,来自该同基因模型的数据支持YTHDF1在结直肠癌中的免疫抑制功能。

图1 在免疫功能正常的小鼠中通过scRNA-seq验证YTH N6-甲基腺苷RNA结合蛋白1(YTHDF1)与结直肠癌(CRC)患者的免疫抑制微环境相关
3、敲除Ythdf1可减少MDSCs,但增加细胞毒性T细胞浸润
为验证在scRNA-seq分析中的发现,作者用流式细胞术测定MC38同基因小鼠肿瘤浸润免疫细胞的组成,证实敲除Ythdf1显著抑制肿瘤的重量和体积(图1C)。并且流式细胞术显示,敲除Ythdf1减少MDSCs,但增加肿瘤中的CD8+ T和CD4+ T细胞(图1H,I)。在MDSCs中,G-MDSCs是主要的亚群,敲除Ythdf1导致G-MDSCs显著减少(图1I)。与MDSCs的免疫抑制功能一致,作者观察到Ythdf1-KO组功能T细胞显著增加,包括IFN-γ+ CD8+ T细胞、颗粒酶B+ CD8+ T细胞和IFN-γ+ CD4+ T细胞(图1H,J)。
接下来,作者在CT26(MSS-CRC)中与Ythdf1-KO进行实验,以验证YTHDF1在调节抗肿瘤免疫中的作用。正如预期的那样,Ythdf1-KO导致CT26同基因小鼠肿瘤体积和重量减小(图2A,B),同时功能T细胞减少和MDSCs积累(图2C,D)。免疫荧光染色证实,Ythdf1基因敲除后,MC38和CT26同基因肿瘤中MDSCs(CD11b+ Gr-1+)的浸润减少(图2E)。总的来说,结直肠癌细胞中Ythdf1的缺失减少MDSCs,增加功能性T细胞的浸润。这些发现与临床数据一致,显示YTHDF1与CD8+ T细胞和IFN-γ相关的特征(图1A-C)。
作者想知道在Ythdf1-KO中减弱的肿瘤形成是否依赖于CD8+ T细胞抗肿瘤免疫。为解决这个问题,作者在MC38同基因模型中使用抗CD8抗体去除CD8+ T细胞。与假设一致,CD8+ T细胞的消耗恢复Ythdf1-KO肿瘤的生长(图2F,G),表明Ythdf1-KO的肿瘤抑制功能至少部分依赖于CD8+ T细胞。这在CT26同基因小鼠中得到证实,表明抗CD8抗体治疗可以挽救Ythdf1-KO组中被抑制的肿瘤生长(图2H,I)。总之,Ythdf1敲除通过诱导CD8+ T细胞依赖性抗肿瘤免疫抑制结直肠癌的生长。

图2 YTH N6-甲基腺苷RNA结合蛋白1(Ythdf1)敲除通过减少髓源性抑制细胞(MDSC)和增加同源肿瘤中的功能性T细胞来诱导抗肿瘤免疫,这种作用被CD8+ T细胞消耗逆转
4、肠道特异性Ythdf1敲入促进小鼠结直肠肿瘤发生并抑制抗肿瘤免疫
为验证YTHDF1在自发性结直肠肿瘤发生中的作用,作者构建肠道特异性的Ythdf1敲入小鼠(Ythdf1loxp/loxp CDX2-cre),并通过AOM/DSS治疗在这些小鼠中启动CRC(图3A),发现在AOM/DSS模型中,Ythdf1的过表达导致结肠肿瘤数量和大小增加(图3C)。流式细胞术显示,与野生型小鼠相比,Ythdf1敲入小鼠结肠肿瘤中MDSC浸润增加,NK、CD4+ T和CD8+ T细胞减少(图3D)。此外,作者发现,通过检测颗粒酶B、INF-γ和TNF-α的表达,Ythdf1敲入降低功能性T细胞的比例(图3E)。
接下来,作者通过建立ApcMin/+ Ythdf1loxp/loxp CDX2-cre小鼠,试图在ApcMin/+驱动的自发性CRC中验证这些结果(图3B)。与此一致的是,肠道特异性敲入Ythdf1的ApcMin/+小鼠的结肠肿瘤明显大于野生型小鼠(图3C)。肿瘤浸润性免疫细胞分析发现ApcMin/+敲入Ythdf1后,肿瘤中NK、CD4+ T和CD8+ T细胞的浸润显著减少,同时MDSCs诱导增加(图3F)。此外,Ythdf1敲入降低ApcMin/+小鼠的颗粒酶B+、INF-γ+或TNF-α+ T细胞(图3G)。总之,这些结果支持YTHDF1培养促进自发性CRC的免疫抑制微环境。
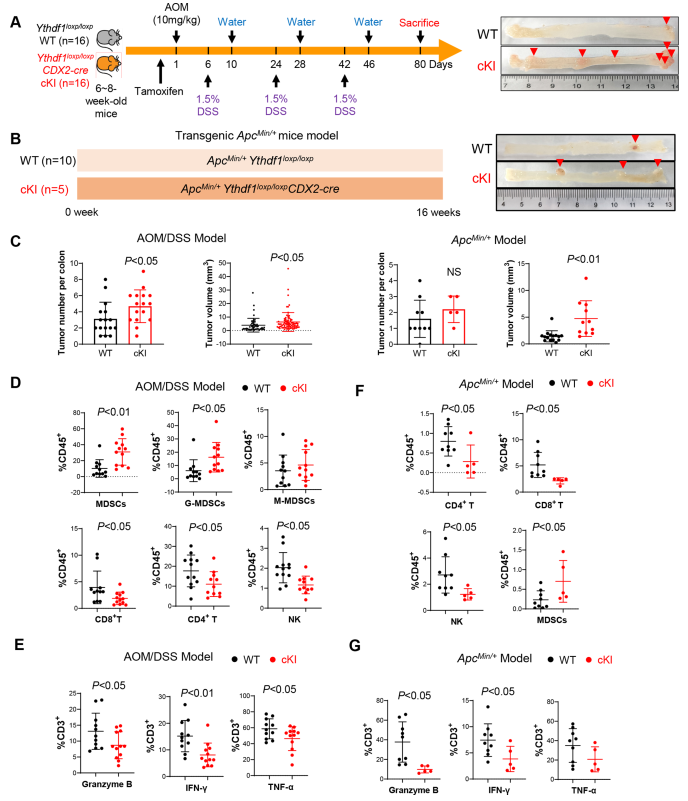
图3 肠道特异性YTH N6-甲基腺苷RNA结合蛋白1(Ythdf1)敲入促进小鼠结肠肿瘤发生并抑制抗肿瘤免疫。
5、通过VNP-siYTHDF1靶向YTHDF1增强CD34+人源化小鼠的抗肿瘤免疫
为证实YTHDF1在调节人抗肿瘤免疫应答中的作用,作者建立CD34+人源化小鼠模型小鼠外周血单个核细胞(PBMCs)含有>20%的人CD45+细胞(图4A)。为在体内靶向YTHDF1,作者开发VNPs来携带针对YTHDF1的siRNA。在肿瘤达到50-100 mm3后,携带人结直肠癌HCT116异种移植物的人源化NSG小鼠接受VNP-siNC或-siYTHDF1治疗(图4B)。与VNP-siNC相比,VNP-siYTHDF1显著抑制肿瘤体积和重量(图4B,C)。进行流式细胞术分析TIME(图4D)。VNP-siYTHDF1减少MDSC浸润,但增加CD4+ T细胞、CD8+ T细胞和NK细胞积聚(图4E)。此外,在接受VNP-siYTHDF1的肿瘤中发现更多的IFN-γ+、TNF-α+和颗粒酶B+ CD8+ T细胞(图4E)。因此,利用VNP-siYTHDF1靶向YTHDF1是增强人源化小鼠抗肿瘤免疫的一种安全有效的手段。

图4 VNP-siYTHDF1增强CD34+人源化小鼠的抗肿瘤免疫
6、YTHDF1促进p65翻译激活TNF/NF-κB信号传导
为确定YTHDF1引发免疫抑制的分子机制,作者在敲除或不敲除YTHDF1的CRC细胞中进行Ribo-seq。Ribo-seq数据显示,YTHDF1的缺失与TNF信号的失活显著相关(图5A)。因此,YTHDF1敲除减少参与TNF信号传导的核糖体保护片段基因丰度(图5A)。因此,YTHDF1可以通过促进蛋白翻译调节TNF/NF-κB信号传导。由于YTHDF1具有m6A阅读器的功能,作者接下来进行m6A免疫沉淀测序(MeRIP-seq)以确定m6A修饰的转录本。通过筛选参与TNF信号传导的mRNA中的m6A峰,作者确定两个靠近p65 mRNA终止密码子的m6A位点(图5B),并通过MeRIP-qPCR验证这一点(图5C)。重要的是,通过RNA免疫沉淀(RIP)测序和抗YTHDF1抗体的RIP-qPCR发现YTHDF1和p65 mRNA之间的直接相互作用(图5D)。因此,p65 mRNA是YTHDF1的直接靶点。作者发现在CT26和MC38细胞中,敲除Ythdf1降低p65蛋白的表达,尤其是核p65的表达,而不影响NF-κB通路的其他调节因子如IKKα和iκBα的表达(图5E)。在人类结直肠癌细胞中获得一致的结果,显示野生型YTHDF1过表达,而非功能失调突变体,p65蛋白表达升高;相反,YTHDF1敲低会减弱人类结直肠癌细胞中的p65蛋白(图5F)。使用抗YTHDF1抗体的RIP-qPCR也证实人结直肠癌细胞中YTHDF1与p65 mRNA之间的直接相互作用(图5G)。接下来,作者试图在体内验证YTHDF1和p65的关联。在敲入Ythdf1的小鼠中,结肠肿瘤中的p65和phospho-p65蛋白均升高(图5H)。总之,YTHDF1促进p65蛋白的表达,在体外和体内激活TNF和NF-κB信号。
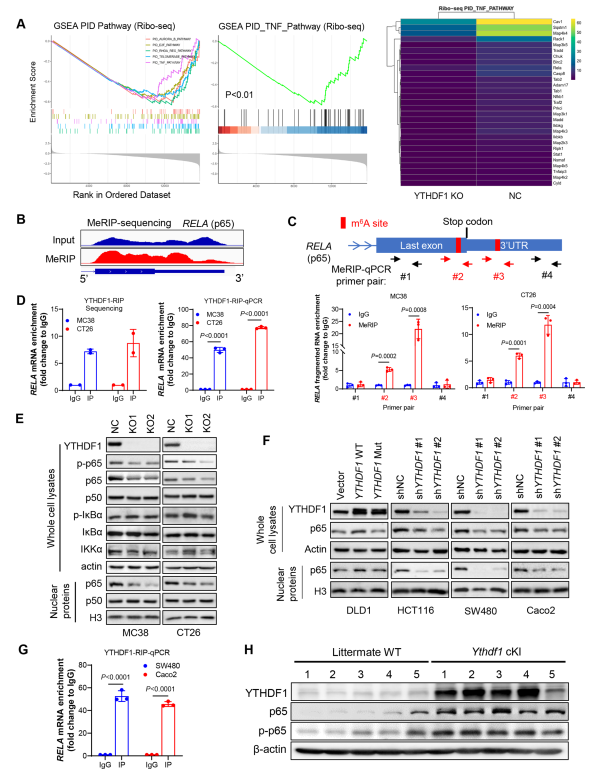
图5 YTH N6-甲基腺苷RNA结合蛋白1(YTHDF1)通过促进RELA (p65) mRNA翻译促进CRC中TNF/NF-kB信号传导
7、YTHDF1通过p65-CXCL1轴促进MDSC迁移
为了解YTHDF1诱导的p65与其免疫抑制之间的联系,作者在CRC细胞、肿瘤裂解物和同源肿瘤小鼠血清的条件培养基中进行细胞因子多重免疫测定,检测23种不同的小鼠细胞因子。在这些细胞因子中,Ythdf1-KO持续降低CXCL1(图6A),并且通过ELISA检测证实CXCL1的这种降低(图6B)。据报道,CXCL1是NF-κB信号传导的转录靶点,它通过与其受体CXCR2的相互作用促进MDSC趋化。鉴于Ythdf1-KO在CRC TIME减少MDSC的浸润,作者想知道YTHDF1是否调节MDSC的迁移。因此,作者进行MDSC体外迁移实验,发现来自野生型CRC细胞的条件培养基增强MDSC的迁移,而这种迁移被Ythdf1的敲除所破坏(图6C)。通过CXCR2抑制剂SB265610阻断CXCL1-CXCR2相互作用,消除对照组和Ythdf1-KO培养上清在介导MDSC迁移方面的差异(图6C)。因此,YTHDF1通过CXCL1/CXCR2轴促进MDSC迁移。接下来,作者想知道YTHDF1是否可以调节Cxcl1 mRNA的表达。正如预期的那样,敲除Ythdf1显著抑制小鼠(图6D)和人CRC细胞系中Cxcl1 mRNA的水平。因此,研究结果支持YTHDF1-p65-CXCL1/CXCR2轴介导CRC中MDSC的迁移。
接下来,作者研究YTHDF1是否影响MDSC功能。从MC38同基因肿瘤中分离CD11b+ Gr-1+ MDSCs,然后与T细胞体外共培养(图6E)。从对照肿瘤中分离的MDSCs抑制T细胞增殖;然而,与来自对照组的MDSCs相比,来自Ythdf1-KO肿瘤的MDSCs对CD8+ T细胞和CD4+ T细胞的增殖抑制活性明显降低(图6F,G)。这些数据验证MDSCs对关键效应细胞(包括CD8+ T细胞和CD4+ T细胞)的免疫抑制功能,并支持表达YTHDF1的CRC招募功能性MDSCs。
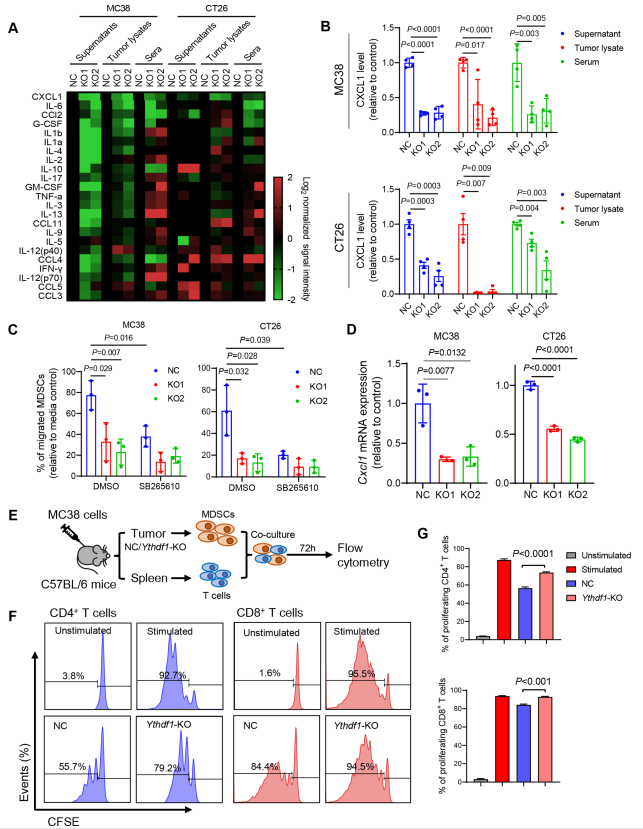
图6 YTH N6-甲基腺苷RNA结合蛋白1(Ythdf1)的缺失通过减少CXCL1的分泌来促进髓源性抑制细胞(MDSCs)的减少。
8、YTHDF1是CRC免疫治疗的潜在治疗靶点
考虑到MDSCs浸润减少已被报道与各种癌症类型的免疫治疗效果增强相关,作者试图测试靶向YTHDF1是否能增强CRC的抗PD1治疗。正如预期的那样,作者发现敲除Ythdf1增强MC38 (MSI-H)同基因肿瘤中抗PD1的功效,延长荷瘤小鼠的生存期(图7A)。作者进一步利用VNPs系统将特异性Ythdf1-siRNA传递到肿瘤中。当MC38同基因肿瘤达到50~100 mm3时,作者用VNP-siYthdf1(或VNP-siNC)和抗PD1(或IgG)治疗小鼠。与VNP-siNC相比,VNP-siYthdf1显著抑制MC38肿瘤生长(图7B,C)。值得注意的是,VNP-siYthdf1与anti-PD1联合使用对肿瘤生长的抑制作用最强(图7B,C)。
基于同基因CT26(MSS CRC)肿瘤模型,作者进一步研究靶向YTHDF1是否可以克服MSS CRC的抗PD1耐药性。因此,将敲除Ythdf1的CT26细胞注射到同基因小鼠体内,并用PD1抗体处理,发现敲除Ythdf1显著增强CT26同基因肿瘤的抗PD1治疗效果,否则这些肿瘤对ICB治疗无反应(图7D,E)。
流式细胞术分析进一步显示,在CT26和MC38同基因模型中,Ythdf1沉默和抗PD1联合使用可显著增加肿瘤浸润的功能性CD8+ T细胞,包括IFN-γ+ CD8+ T细胞和颗粒酶B+ CD8+ T细胞(图7F,G)。此外,联合治疗显著减少MDSCs的积累,同时诱导CD4+ T细胞和CD8+ T细胞(图7G)。因此,靶向YTHDF1不仅可以增强ICB在MSI-H型CRC中的治疗效果,还可以通过抑制MDSCs的募集和改善CD8+ T细胞的功能来克服MSS型CRC中的ICB耐药性。
在这里,作者发现在癌细胞中消耗Ythdf1会抑制肿瘤内的MDSCs,并减轻MDSCs对效应细胞的免疫抑制功能。MDSCs和T细胞共培养研究表明,与对照肿瘤相比,来自Ythdf1-KO肿瘤的MDSCs抑制CD8+ T细胞和CD4+ T细胞增殖的能力受损。因此,研究结果支持YTHDF1介导功能性MSDCs的募集,这抑制CRC中效应T细胞的功能和增殖,导致免疫监视功能受损(图7H)。

图7 靶向YTH N6-甲基腺苷RNA结合蛋白1(YTHDF1)增强抗PD1阻断治疗在微卫星不稳定性高(MSI-H)和微卫星稳定型结直肠癌(CRC)中的应用
综上所述,YTHDF1在结直肠癌中的表达通过激活m6 A-p65-CXCL1轴募集免疫抑制性MDSCs抑制T细胞,从而促进结直肠癌的发生。靶向YTHDF1加抗PD1治疗对结直肠癌有良好的抗肿瘤疗效,证实YTHDF1是结直肠癌的潜在治疗靶点。
实验方法
人源化小鼠模型,TCGA数据分析,CRISPR/Cas9敲除,稳定细胞系过表达,同基因小鼠模型,RT-qPCR,免疫组织化学,免疫荧光,Western blot,酶联免疫测定(ELISA),RNA干扰,囊泡样plga基纳米颗粒(VNP)配方,流式细胞术分析,单细胞RNA测序,MDSC分离及迁移试验,细胞因子多重免疫测定,核糖体测序,MeRIP测序和MeRIP-qPCR,RIP-测序或RIP-qPCR,肝肾功能指标测定
参考文献
Bao Y, Zhai J, Chen H, et al. Targeting m6A reader YTHDF1 augments antitumour immunity and boosts anti-PD-1 efficacy in colorectal cancer. Gut. 2023; 72 (8): 1497-1509. doi: 10.1136/gutjnl-2022-328845