






SIRT6调节的巨噬细胞胞葬作用表观遗传控制糖尿病牙周炎的炎症消退
糖尿病加剧了牙周炎的患病率和严重程度,导致严重的牙周破坏并最终导致牙齿脱落。炎症的延迟消退是糖尿病牙周炎(DP)发病机制的主要因素,但这种不平衡的免疫稳态的潜在机制尚不清楚。下图为23年巨噬细胞胞葬中标题目:
巨噬细胞中SIRT6作为组蛋白去乙酰化酶的不足导致人类和小鼠DP中未解决的炎症和加重的牙周炎,并伴有凋亡性中性粒细胞(AN)的积累和中性粒细胞胞外陷阱的增加。在机制上,验证了巨噬细胞接受高糖刺激导致SIRT6-miR-216/217轴紊乱,通过直接靶向DEL-1/CD36轴触发AN的胞葬受阻。此外,我们证明了SIRT6对MIR217HG转录的抑制作用,并鉴定了微处理器的作用,即SIRT6通过hnRNPA2B1、DGCR8和Drosha复合体在表观遗传学上阻碍了初级miR-216/217的剪接。该研究于23年1月发表于《Theranostics》,IF:12.4。
技术路线:

1.嗜中性粒细胞和巨噬细胞在人DP中的特征性分布
为了探索糖尿病相关炎症性疾病中细胞群组成和调节细胞通路的差异,使用单细胞测序分析了来自健康非糖尿病和糖尿病足溃疡非治愈者(DFU-Non-healer)的足部皮肤样本GSE165816 .GO富集分析结果显示,炎症和细胞凋亡相关通路被激活(图1A和B)。在鉴定的炎症细胞中,研究专注于增加巨噬细胞浸润,并通过GO分析和UMAP分析进一步鉴定巨噬细胞簇的细胞群。UMAP分析显示巨噬细胞异常极化(图1C)。GO富集分析显示,与非DM对照巨噬细胞相比,糖尿病巨噬细胞在免疫反应、吞噬作用、迁移能力和葡萄糖代谢等相关途径中出现异常(图1D和E)。这些结果表明,糖尿病创面愈合过程中的异常炎症反应可能是由细胞凋亡相关通路的大量激活和巨噬细胞的异常吞噬能力引起的。为了进一步探讨DP炎症持续升高是否与巨噬细胞吞噬能力异常有关,我们收集了患有或不患有糖尿病的牙周炎患者的牙龈样本(图1F)。因此,我们首先在牙龈中检测到中性粒细胞,结果显示DP中性粒细胞浸润(CD15)细胞数量高于CP组(1G和L)。采用连续切片牙龈组织进行TUNEL染色,结果显示DP的牙龈组织中有大量的凋亡细胞,而CP中几乎没有可见的凋亡细胞(图1H和M)。此外,通过切片进行比较观察,发现大多数凋亡细胞是CD15中性粒细胞(图1G和H)。随后使用MPO和H3cit标记NETs,发现DP中中性粒细胞的过度积累和延迟去除导致了大量NETs的形成(图1I和N),从而加重了糖尿病患者的炎症并延迟了炎症的消退。为进一步探究未及时清除的凋亡中性粒细胞是否为巨噬细胞浸润异常所导致的,接下来对巨噬细胞极化进行免疫荧光染色。巨噬细胞(CD68)在DP牙龈中的浸润率远高于CP(图1O)。DP组CD68、CD86阳性M1巨噬细胞浸润率高于CP组(图1J和P),CD68 CD206阳性M2极化巨噬细胞中DP组和CP组无差异(图1K和Q)。然而,DP组的M2/M1比值明显低于CP组(图1R)综上所述,凋亡中性粒细胞和NETs的过度积累和延迟清除,伴有巨噬细胞浸润增加和巨噬细胞极化破坏,无疑加重了DP炎症。
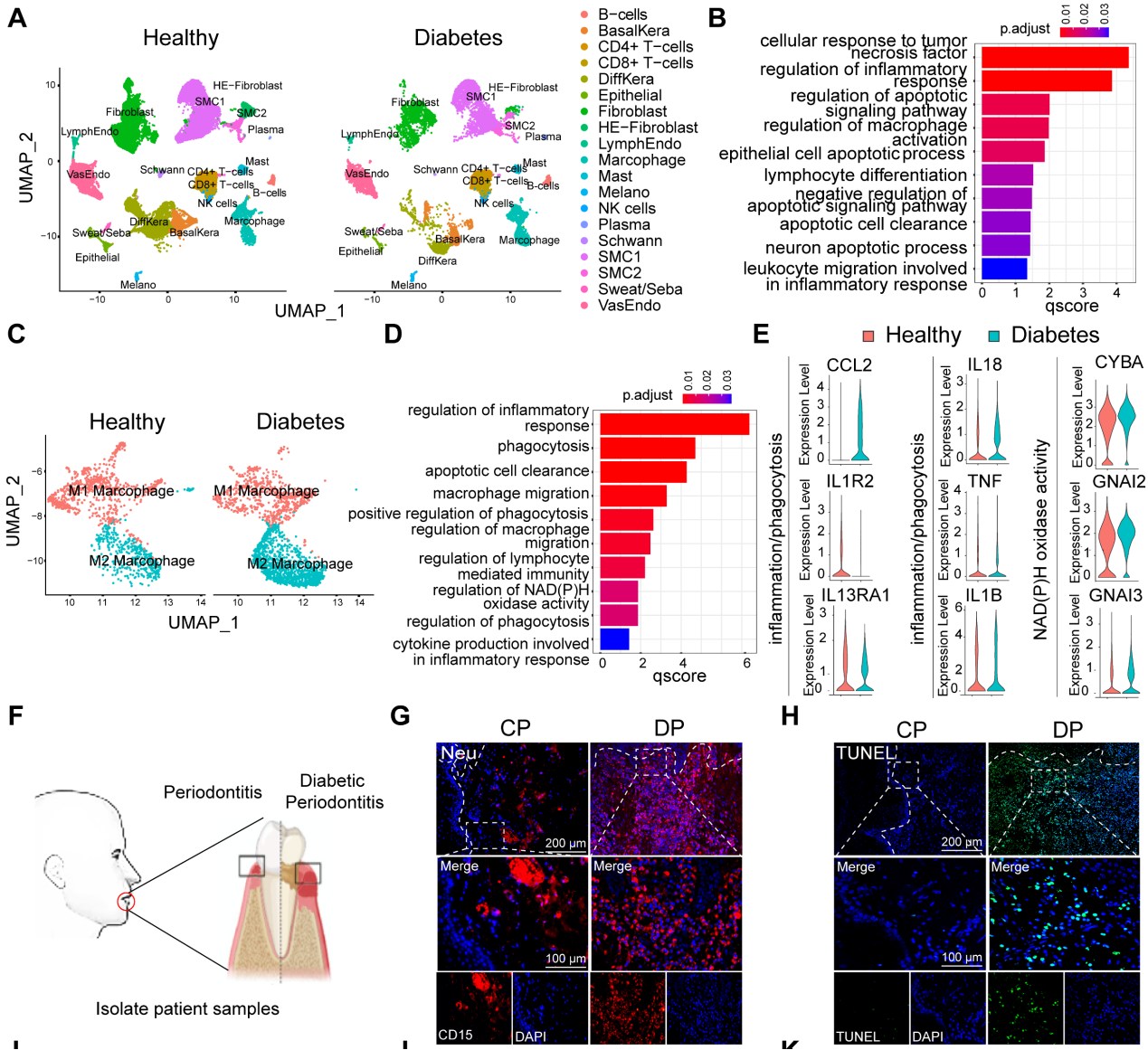
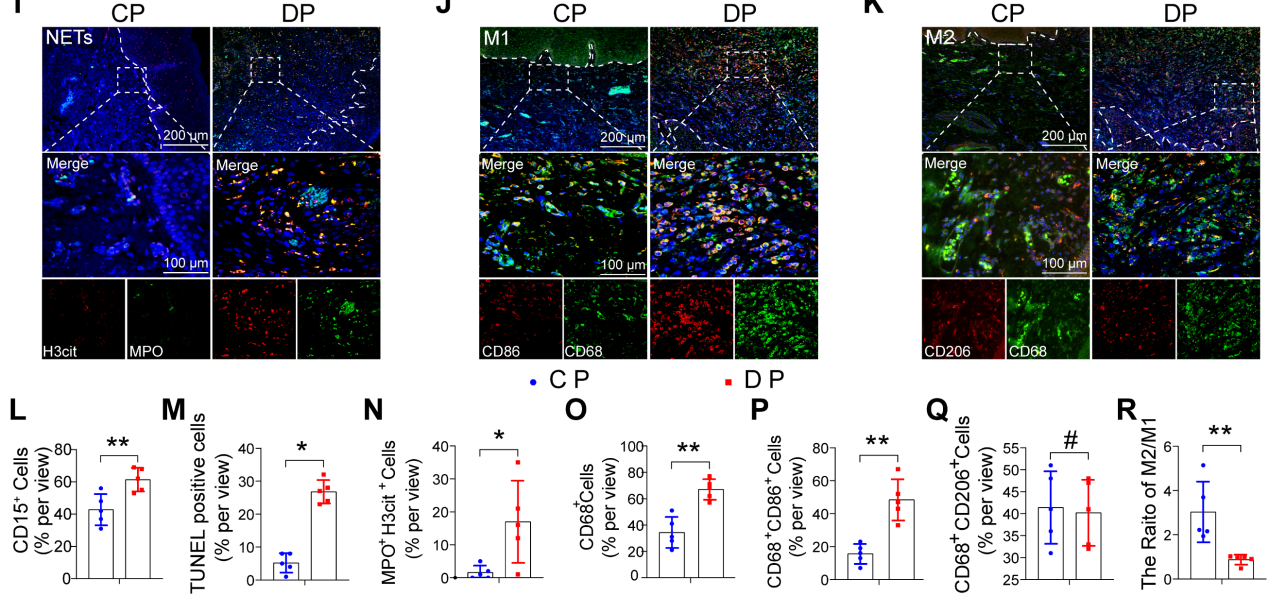 图1嗜中性粒细胞和巨噬细胞在人DP中的特征性分布
图1嗜中性粒细胞和巨噬细胞在人DP中的特征性分布
2.功能失调的中性粒细胞和巨噬细胞会加重小鼠 DP 的炎症损伤并损害炎症消退
为了探究糖尿病对牙周炎进展和炎症消退的影响,我们使用了两种经典的结扎诱导牙周炎(LIP)小鼠模型,即牙周炎模型和牙周炎消退模型。在牙周炎模型中进行结扎过程长达14天,而牙周炎消退模型中的小鼠结扎7天,然后去除结扎以模拟炎症消退的过程再进行7天(图2A)进行显微CT和组织形态学分析以观察牙槽骨丢失和炎症。在牙周炎和牙周炎消退模型中,糖尿病小鼠从CEJ到ABC的距离均显着增加(图2B和I)。在牙周炎和牙周炎消退模型中,与CP组相比,糖尿病小鼠表现出更多的破骨细胞活性,TRAP阳性破骨细胞表面发生率更高(图2C和J)。此外,在DP的牙周中观察到大量的中性粒细胞浸润以及MPO和H3cit NET(图2D、K、E、L)。TUNEL染色还显示DP牙周内有大量凋亡细胞,而CP组未观察到可见的凋亡细胞(图2F和M)。DP组F4/80巨噬细胞浸润数大于CP组(图2N)具体来说,F4/80+ CD86+ M1巨噬细胞的数量(图2G和O)和F4 / 80 CD206 M2巨噬细胞(图2H和P)显著高于CP组。然而,在LIP模型的分辨率中,与CP组相比,DP中M2 / M1巨噬细胞的比率也显着降低(图2Q)总的来说,牙周组织中凋亡性粒细胞和NETosis的积累加速了牙周炎的进展,而凋亡性中性粒细胞和NET的延迟清除阻碍了小鼠DP炎症的消退。
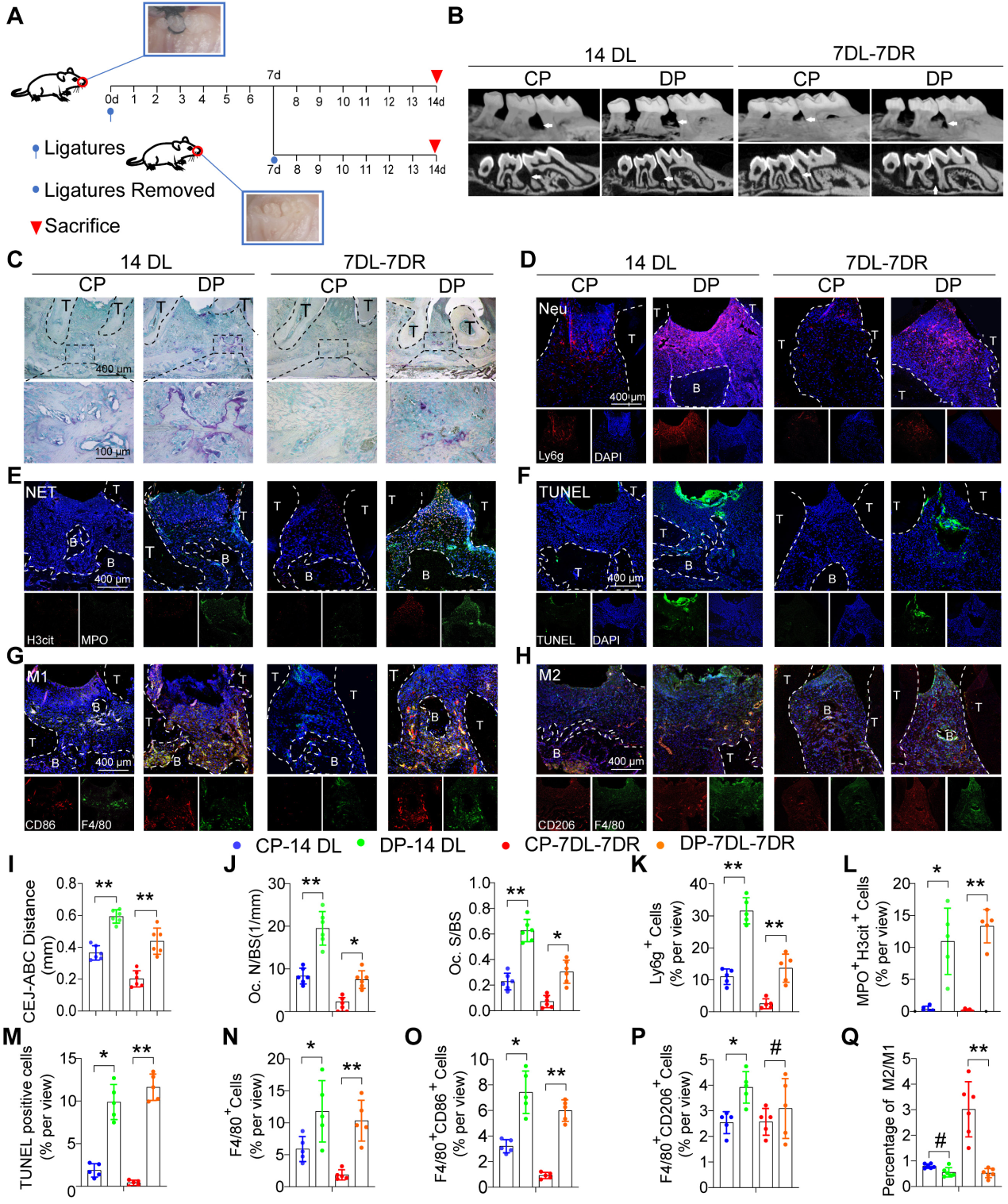
图2功能失调的中性粒细胞和巨噬细胞会加重小鼠 DP 的炎症损伤并损害炎症消退
3.SIRT6 在高糖条件下显着调节巨噬细胞胞吐作用
高糖(HG)刺激后巨噬细胞吞噬凋亡中性粒细胞的能力下降(图3A)。鉴于DM巨噬细胞团团的GO富集分析显示NAD+代谢异常(图1D, E),我们进一步检查发现HG刺激可导致NAD+/NADH代谢异常(图3B)。Sirtuins (SIRTs)是一个依赖NAD+的组蛋白去乙酰化酶家族,在葡萄糖稳态、炎症、基因组稳定性和DNA修复中发挥作用。因此,我们确定并重点关注SIRT6,因为它在HG条件下明显下调,在48 h和72 h时最为明显(图3C)。为了进一步研究SIRT6在DP巨噬细胞中的可能功能,CD68(巨噬细胞标志物)和SIRT6共染色显示,与CP相比,DP巨噬细胞中SIRT6的表达显著降低(图3D)。SIRT6抑制剂显著降低了巨噬细胞进行胞葬的能力,而SIRT6过表达则提高了其吞噬凋亡中性粒细胞的能力(图3E, F)。与胞葬相比,凋亡细胞增加了巨噬细胞的迁移能力。综上所述,高糖刺激导致巨噬细胞SIRT6的低表达,从而损害了胞葬,胞葬通过减少死细胞的DAMP释放和维持生物稳态来抑制炎症。然而,当糖尿病患者巨噬细胞的胞浆功能受损时,通过损伤相关的分子识别模型,促炎溶解细胞死亡,包括NETosis和胞葬,导致巨噬细胞浸润增加,打破了M1和M2分化的平衡。
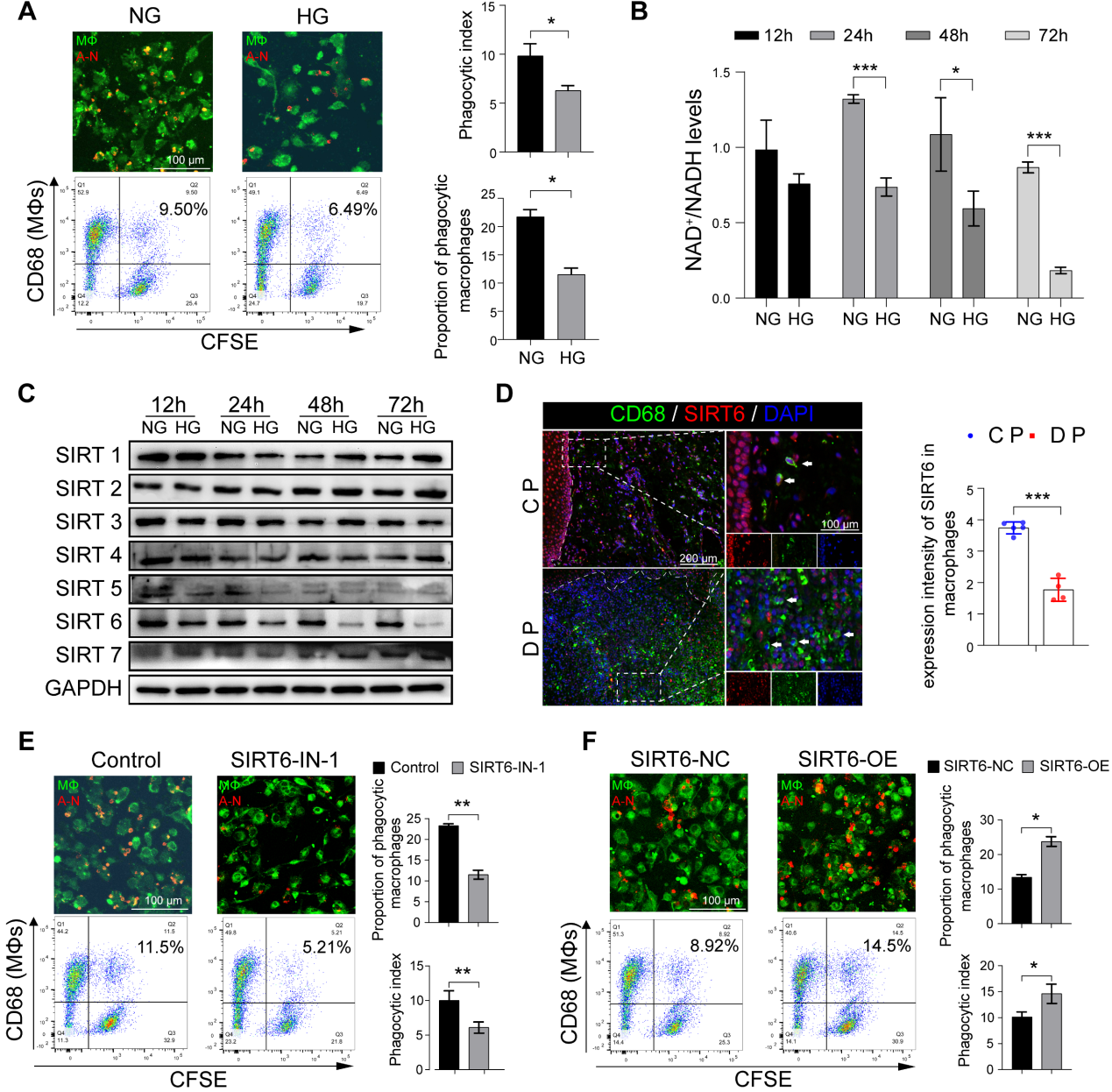
图3 SIRT6 在高糖条件下显着调节巨噬细胞胞吐作用
4.髓细胞特异性SIRT6基因敲除会加重牙周炎并损害炎症消退
为了探索SIRT6介导的巨噬细胞胞葬在牙周炎中的作用,我们将LysM-Cre小鼠与SIRT6flox/flox小鼠杂交产生mS6KO小鼠。与野生型小鼠骨髓巨噬细胞(BMMs)相比,mS6KO骨髓巨噬细胞吞噬凋亡中性粒细胞的能力下降(图4A)。采用2月龄mS6KO小鼠及其WT窝代建立LIP模型和LIP分辨率模型。与WT小鼠相比,CEJ到ABC的距离(图4B, I)和捕获阳性的破骨细胞明显增加(图4C, J)。为了研究巨噬细胞胞葬功能受损是否与mS6KO小鼠的持续性骨质流失有关,通过TUNEL染色评估mS6KO小鼠和WT小鼠牙周组织中凋亡细胞的数量(图4D,结果显示,与同窝野生型小鼠相比,mS6KO小鼠牙周组织中Ly6g+细胞和凋亡细胞的数量显著增加(图4E, L), SIRT6敲除促进了mS6KO小鼠牙周组织中NETosis(图4F, M), F4/80+ CD86+ M1巨噬细胞(图4G, O)和F4/80+ CD206+ M2巨噬细胞(图4H, P)。与WT小鼠相比,mS6KO小鼠牙周组织中抗炎的F4/80+ CD206+ M2巨噬细胞的百分比较低(图4Q)。综上所述,这些发现表明SIRT6介导的巨噬细胞胞葬与牙周破坏和炎症的消退有关。
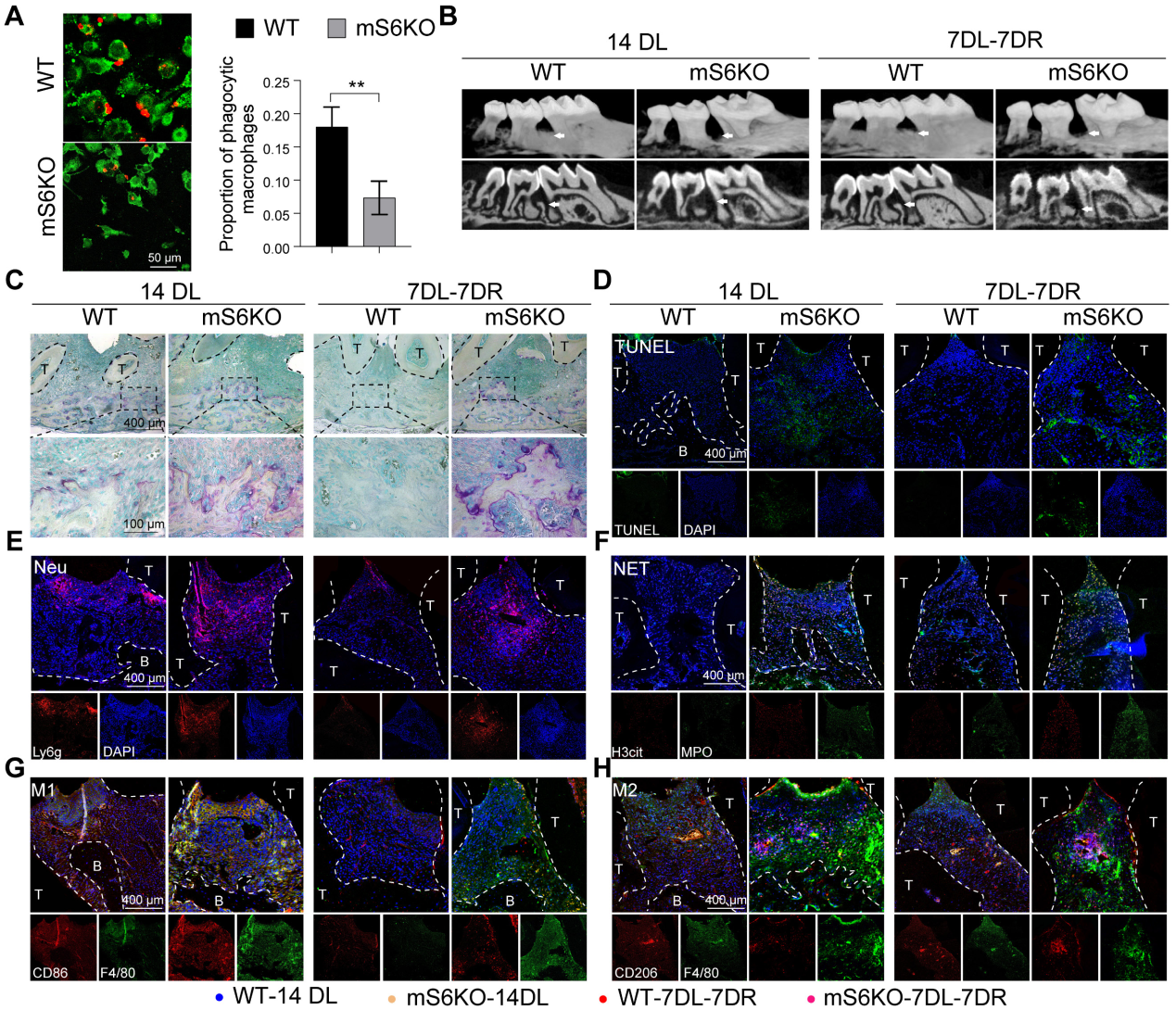
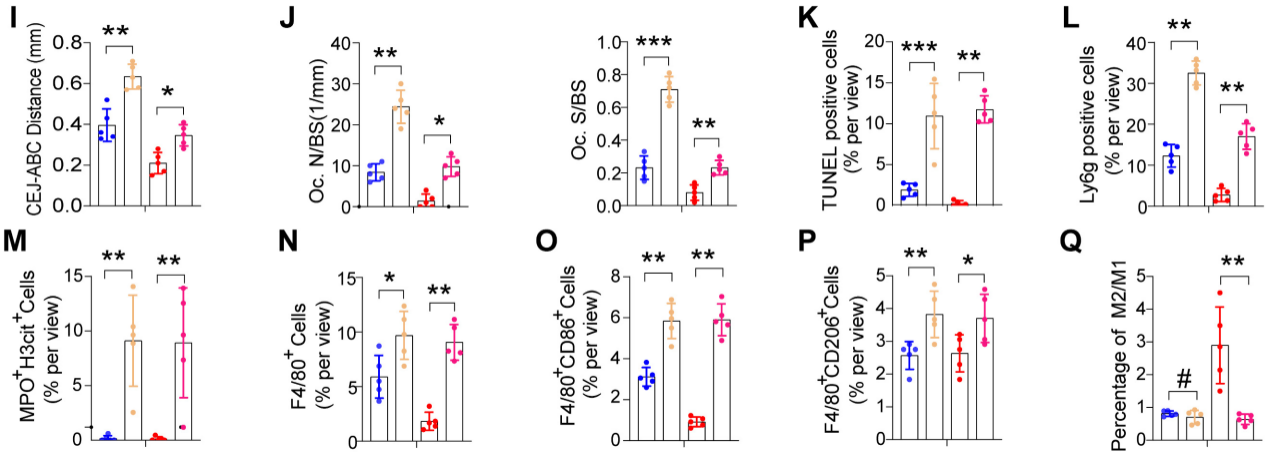
图4髓细胞特异性SIRT6基因敲除会加重牙周炎并损害炎症消退
5.SIRT6通过H3K56ac抑制miR-216a-5p-216b-5p-217簇的转录
为了深入了解SIRT6调控巨噬细胞胞葬的详细机制,我们进一步评估了SIRT6对巨噬细胞中microRNA表达的影响。从SIRT6抑制后的巨噬细胞中分离RNA进行miRNA PCR微阵列分析,结果显示miR-216a-5p-216b-5p-217簇明显增加,SIRT6表达下调(图5A-C)。数据进一步显示SIRT6抑制和HG条件下miR-216a-5p-216b-5p-217簇表达增加(图5D和F),SIRT6过表达导致miR-216a-5p-216b-5p-217簇表达降低(图5E)。为了进一步阐明SIRT6如何抑制miR-216a-5p-216b-5p-217簇表达,通过ChIP-qPCR检测了巨噬细胞中SIRT6宿主基因MIR217HG启动子的组蛋白修饰。SIRT6作为一种转录共抑制因子,通过催化位点9和56乙酰化的组蛋白H3赖氨酸残基(H3K9ac和H3K56ac)的去乙酰化。HG处理仅显著上调H3K56ac的表达(图5H)。使用覆盖miR-216a-5p-216b-5p-217启动子的七对引物进行检测,结果显示HG处理增加了miR-216a-5p-216b-5p-217集群启动子的H3K56ac水平(图5I)。我们还进一步证实了SIRT6抑制和HG条件下pri-miR-217簇表达增加(图5J和L),SIRT6过表达后pri-miR-217簇表达下调(图5K)。值得注意的是,来自mS6KO小鼠的bmm显示出增加的pri-miR-217和miR-216a-5p-216b-5p-217簇表达(图5G和M)。
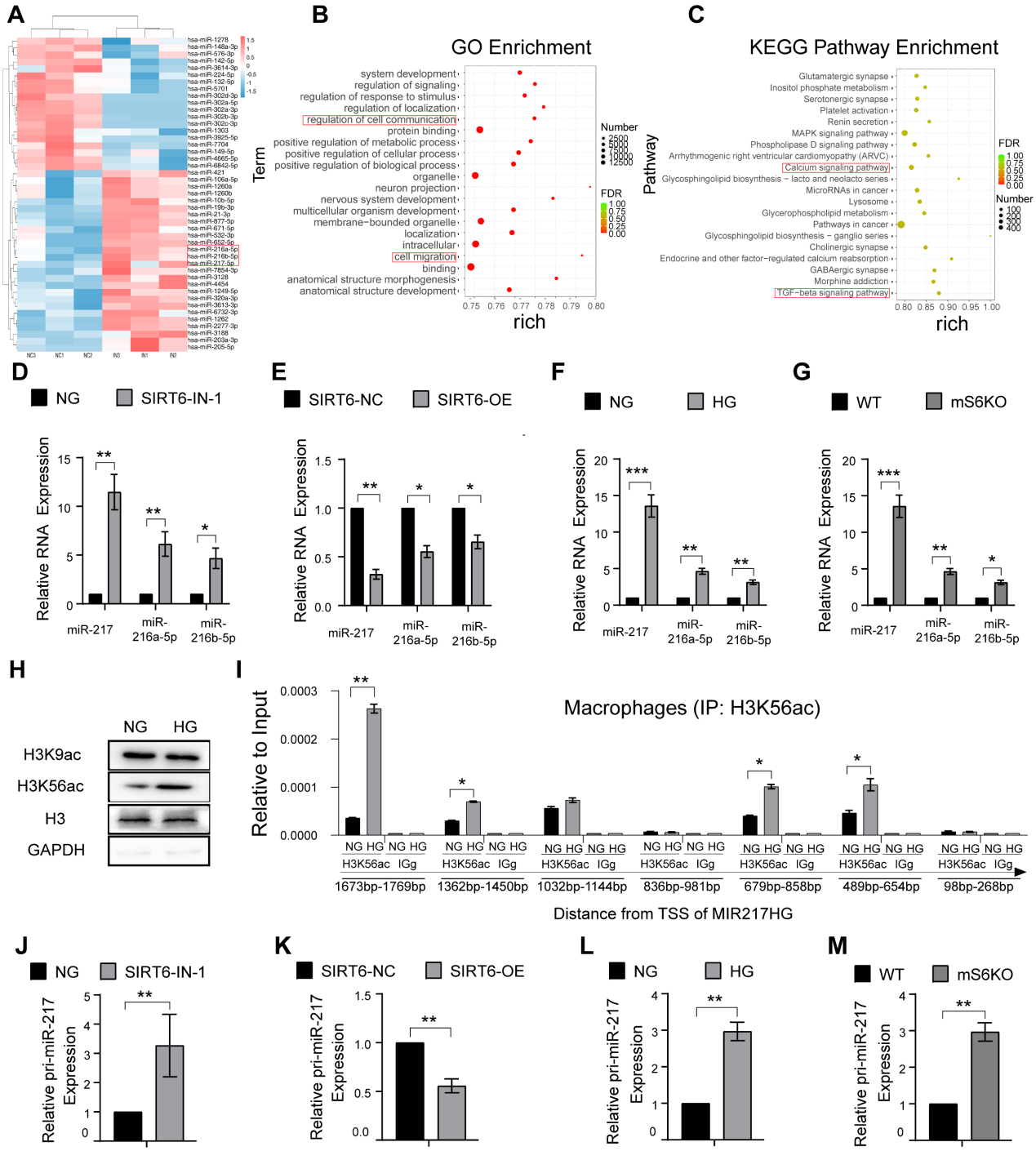
图5 SIRT6通过H3K56ac抑制miR-216a-5p-216b-5p-217簇的转录
6.SIRT6 通过“非经典”微处理器复合体抑制 miR-216a-5p-216b-5p-217 簇成熟
有研究发现,葡萄糖代谢影响Drosha蛋白的表达,葡萄糖剥夺促进Drosha表达,HG刺激抑制Drosha表达。同样,我们的结果显示HG刺激可以抑制巨噬细胞中Drosha蛋白的表达(图6A)。为了进一步探索HG条件下miR216a-5p-216b-5p-217簇成熟的机制,我们设计了一种特异性的生物素标记的pri-miR-217探针,在巨噬细胞中进行RNA下拉实验,银染显示与pri-miR-217结合的几个蛋白带富集(图6B)。同时,蛋白质谱分析显示hnRNPA2B1在识别蛋白列表中排名靠前,未观察到Drosha和DGCR8。RIP实验显示,与IgG相比,hnRNPA2B1抗体降低了更多的pri-miR-217(图6C)。据报道,hnRNPA2B1与microRNA微处理器复合物蛋白DGCR8相互作用,通过结合m6A标记初级miRNA转录物来切割初级miRNA。内源性hnRNPA2B1蛋白与巨噬细胞中DGCR8和Drosha的共免疫沉淀(Co-IP)表明它们之间存在直接的物理相互作用,hnRNPA2B1与DGCR8结合,而未观察到Drosha(图6F)。我们进一步确定DGCR8可以与Drosha结合,这意味着hnRNPA2B1与DGCR8结合,招募Drosha形成微处理器复合物,“非规范”微处理器复合物识别并切割pri-miR-217(图6E)。Co-IP还显示SIRT6抑制促进了“非规范”微处理器复合物的形成,SIRT6过表达减少了微处理器复合物(图6G)。此外,我们进一步发现,高葡萄糖不仅增加了与pri-miR-217结合的hnRNPA2B1(图6D)。Chip的结果还显示,HG处理增加了hnRNPA2B1启动子的H3K56ac(图6H)。为了确定HG是否以hnRNPA2B1依赖的方式影响pri-miR-217的加工,我们对巨噬细胞进行了hnRNPA2B1敲低,发现成熟miRNA的水平下降,而其pri-miR-217的水平上升(图6I, J)。FISH染色显示HG刺激导致细胞核中pri-miR-217的减少,以及敲低hnRNPA2B1后细胞核中pri-miR-217的积累(图6K)。这些数据表明SIRT6不仅可以促进pri-miR-217的转录,还可以通过促进HG条件下“非规范”微处理器复合物的形成来促进miR216a-5p-216b-5p-217簇的成熟。
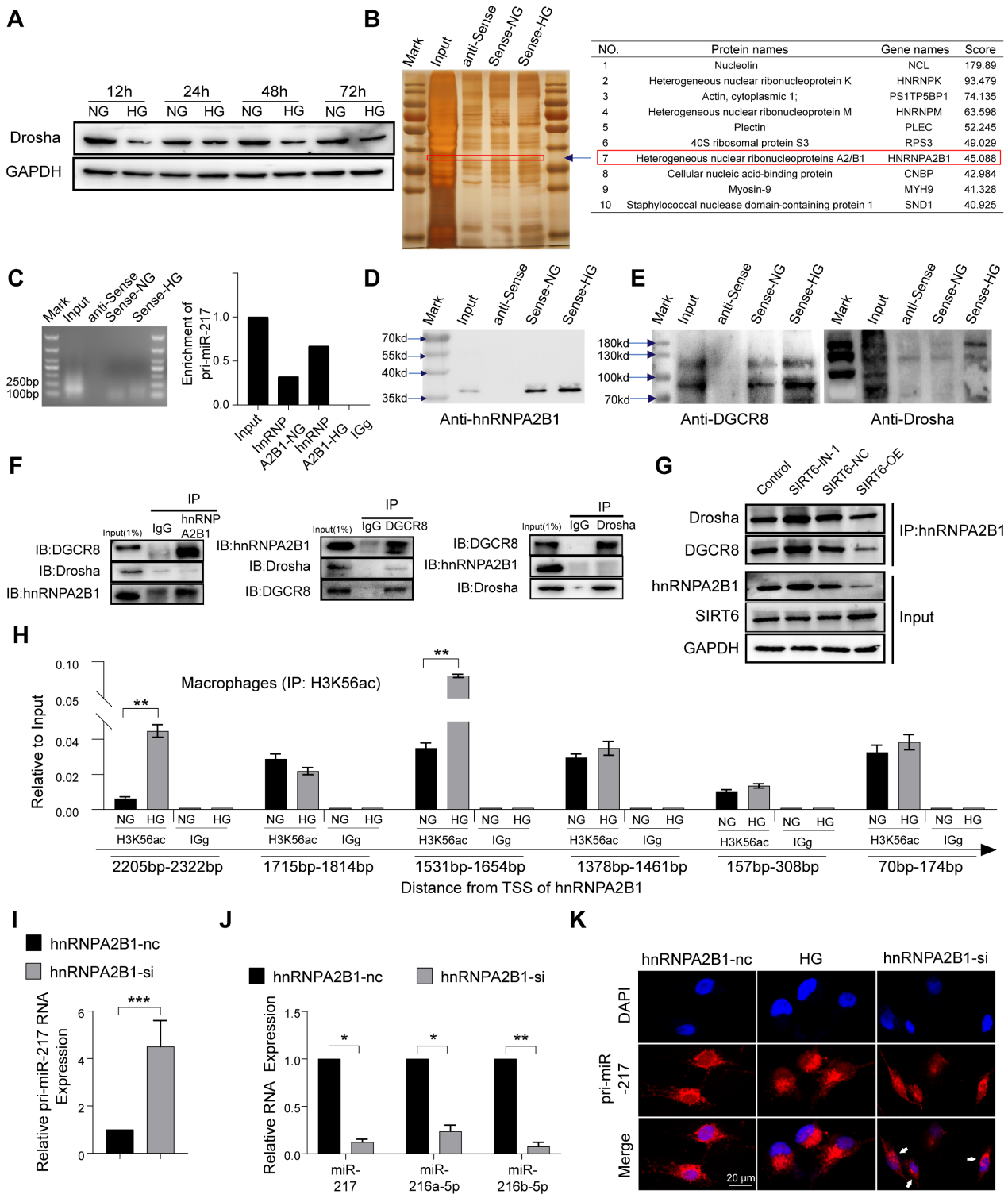
图6 SIRT6 通过“非经典”微处理器复合体抑制 miR-216a-5p-216b-5p-217 簇成熟
7.miR-216a-5p-216b-5p-217簇通过靶向DEL-1和CD36负性调节巨噬细胞的胞葬
DEL-1和CD36已被确定为巨噬细胞胞葬和炎症清除的关键调节分子。我们假设SIRT6通过miR-216/217簇/ DEL-1 CD36轴调节巨噬细胞的胞葬和炎症的消退。生物信息学算法(miRwalk)的结果表明,miR-216a-5p-216b-5p-217簇与DEL-1和CD36 mRNA的3'UTR结合(图7A)。随后,双荧光素酶报告基因实验显示,miR216a-5p216b-5p-217簇的共转染降低了WT-DEL-1-3'UTR和WT-CD36-3'UTR的荧光素酶活性,而Mut-DEL-1-3'UTR和Mut-CD36 -3'UTR的荧光素酶活性不受影响(图7B)。此外,Western blot分析显示,在miR216a-5p-216b-5p-217 mimic处理的巨噬细胞中,DEL-1和CD36表达降低,而在miR-216b5p-217 inhibitor处理的巨噬细胞中,DEL-1和CD36表达升高(图7C)。免疫荧光也证实了DEL-1和CD36在糖尿病牙周炎中的低表达(图7D)。miR216a-5p-216b-5p-217簇的过表达降低了巨噬细胞的吞噬能力(图7E),在这三种micRNA中,miR-217对巨噬细胞的吞噬作用影响最大。HG环境下miR-217敲低可以像重组DEL-1蛋白一样恢复巨噬细胞的胞葬(图7F)。总体而言,我们的研究结果支持SIRT6通过促进pri-miR-217转录和pri-miR“非规范”微处理器复合物的形成负调控miR216a-5p/ 216b-5p-217簇表达的假设。该microRNA簇通过靶向关键的胞葬分子DEL-1和CD36抑制巨噬细胞的胞葬,导致巨噬细胞介导的炎症消退功能失调。
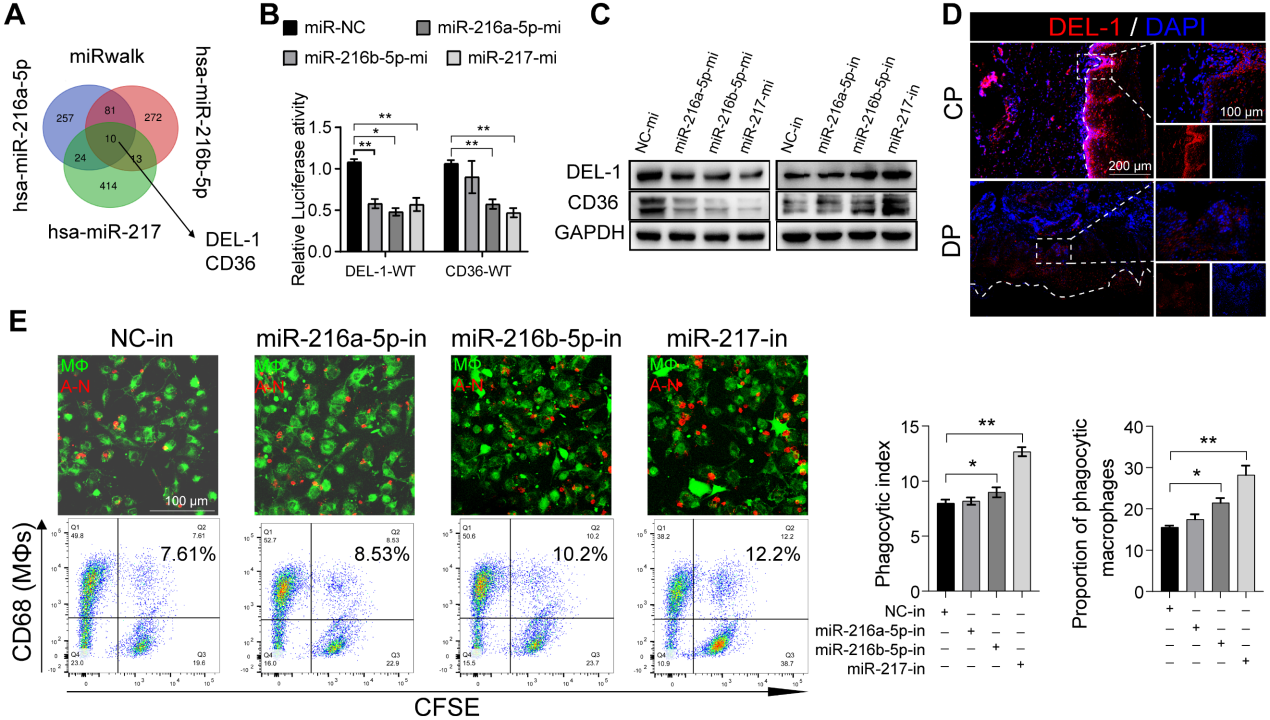
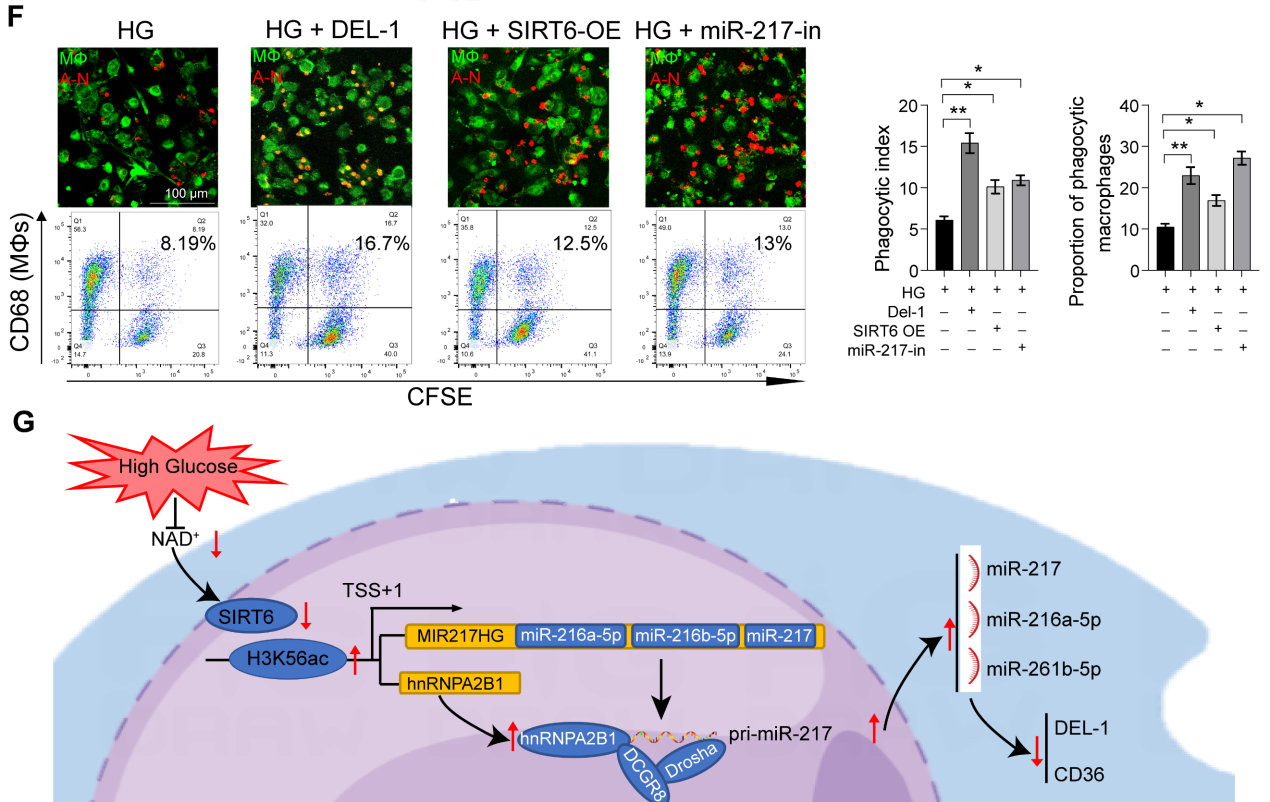
图7miR-216a-5p-216b-5p-217簇通过靶向DEL-1和CD36负性调节巨噬细胞的胞葬
8.局部给药antagomir-217促进小鼠DP炎症的消退
根据上述功能获得和功能丧失研究中的吞噬指数观察,miR-217表现出更好的表型,并被选择用于体内干预治疗。我们在结扎丝后第3、6、9和12天立即将miR-217特异性拮抗剂或scramble (NC)-miR注射到糖尿病小鼠的牙周组织中(图8A)。在第7天解除结扎后,安他哥米-217注射液显著改善了糖尿病小鼠牙周炎症和骨质丢失(图8B、C、G、H)。注射安他哥米-217后,糖尿病小鼠牙周组织中DEL-1的表达显著增加(图8D和I)。注射安他哥米-217后,糖尿病小鼠牙周组织中Ly6g+中性粒细胞的数量显著减少(图8E和J)。注射安塔戈米尔-217后,MPO+ H3cit+ NET(图8L)和凋亡细胞的形成明显减少(图8F和K)。安塔戈米尔-217治疗组M2抗炎巨噬细胞比例较高(图8O),而对照组大部分巨噬细胞为M1巨噬细胞(图8M, N, P)。总而言之,这些结果表明,通过沉默miR-217抑制SIRT6 / miR216a-5p-216b-5p-217簇/ DEL-1和CD36调节轴可促进巨噬细胞的胞葬以清除凋亡细胞,这意味着该调节轴具有作为糖尿病炎症解决靶点的潜力。
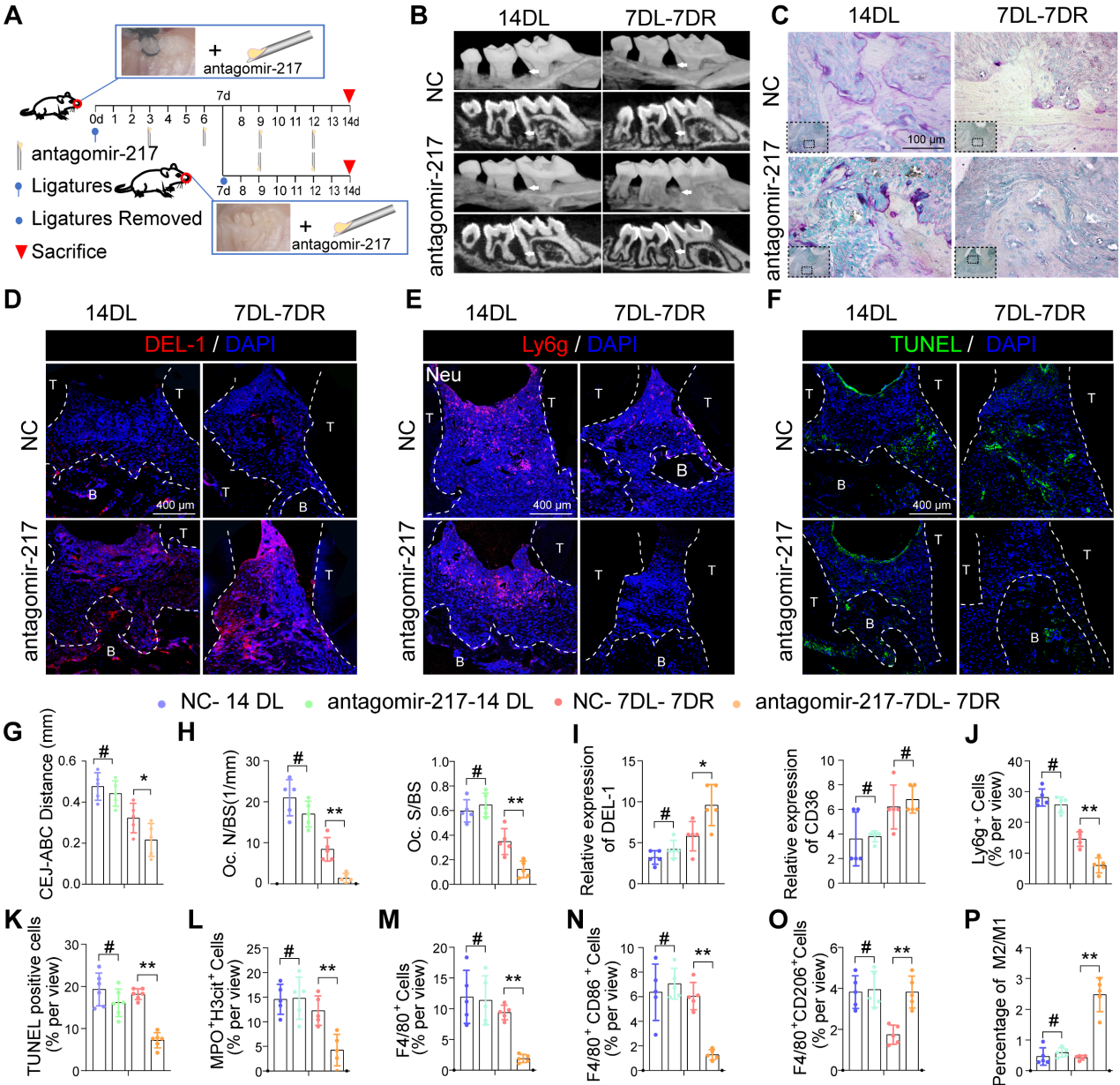
图8局部给药antagomir-217促进小鼠DP炎症的消退
结论:
综上所述,作者揭示了SIRT6-miR-216/217轴在糖尿病背景下巨噬细胞胞葬中的重要作用,并概述了通过抑制miR-217来改善炎症消退和牙周组织修复的方法。这一策略可能与糖尿病和其他慢性炎症性疾病的管理相关。
实验方法:
小鼠牙周炎和消退造模、苏木精和伊红、Masson 染色、免疫荧光染色、流式检测凋亡细胞的吞噬作用、RNA pull-down 、RIP、ChIP、FISH、Co-IP、双荧光素酶报告基因。
参考文献:
Li B, Xin Z, Gao S, et al. SIRT6-regulated macrophage efferocytosis epigenetically controls inflammation resolution of diabetic periodontitis. Theranostics. 2023;13(1):231-249. Published 2023 Jan 1.