






METTL16的乳酸化通过FDX1 mRNA的m6A修饰促进铜死亡
m6A是RNA的一种表观遗传修饰,将这种化学标记添加到RNA分子中,通过影响RNA分子的命运来调节基因表达。这种转录后RNA修饰是可逆的,由甲基转移酶“writers”和去甲基化酶“erasers”调节。m6A修饰的RNA的命运取决于识别和结合它们的不同“readers”的功能。由于m6A甲基化修饰在调节癌症进展中的重要作用。下图为2023年m6A修饰的中标项目:
胃癌是最常见的恶性肿瘤之一,是全球癌症相关死亡的第四大原因。幽门螺杆菌感染、过量摄入盐和硝酸盐以及其他环境风险因素对胃癌的发生有很大影响。此外,遗传改变和宿主因素也与GC的发生和发展有关。尽管手术和辅助治疗方法有所改善,但晚期GC患者的预后仍然较差,5年生存率<25%。更好地了解这种致命疾病的潜在机制并制定更有效的靶向治疗策略对于改善胃食癌患者的临床结果是必要的。该文章于2023年10月发表在《nature communication》,IF:16.6
技术路线: 
研究结果:
1.胃癌中铜含量过高,与肿瘤进展有关
铜死亡是由铜浓度过高引起的,但肿瘤中铜死亡的调控机制仍有待探索。由于铜主要通过胃和小肠上部吸收,胃肠道肿瘤适合研究铜增生的发生和发展。我们分析了48对胃癌组织和邻近组织中的铜浓度,发现胃癌组织中的铜浓度高于正常胃组织(图1A, P < 0.01)。进一步分析发现,III期胃癌患者的相对铜含量高于I期和II期胃癌患者(图1B, P < 0.05),表明铜含量与肿瘤进展相关。此外,铜含量与GC患者的总生存期(OS)和无病生存期(DFS)呈负相关(图1C, d;log-rank P < 0.05)。同时,铜含量与细胞增殖指标Ki-67的表达呈正相关(图1E)。同样,铜含量与患者血清标本中血小板/淋巴细胞比率和中性粒细胞/淋巴细胞比率呈正相关,这两者都是GC浸润性恶性肿瘤炎症相关的预后生物标志物(图1F, G)。这些结果表明,高铜含量参与了GC的进展和发展。此外,我们还检测了不同GC类型中铜浓度的差异,特别是在恶性肿瘤类型中。有趣的是,我们发现粘液腺癌中的铜浓度高于整个胃肿瘤(图1H)。在粘液腺癌中,铜含量与浸润性指标如淋巴结转移(图1I)和血管浸润(图1J)呈正相关。黏液性腺癌是一种罕见的组织学亚型,与非黏液性腺癌相比,黏液性腺癌的分期更晚期,5年OS和DFS更差。粘液腺癌是一种难治性胃癌,具有放化疗耐药的特点,迫切需要更有效的治疗方法。最近,研究人员发现,高细胞内铜浓度触发还原酶FDX1将Cu2+还原为毒性更强的Cu+形式,导致铜死亡。我们发现,与正常胃组织相比,GC中的FDX1蛋白水平更高(图1K)。由于GC中FDX1蛋白水平和铜含量较高,可能更容易引发铜沉积。它为胃癌提供了一种潜在的治疗策略,特别是对恶性肿瘤-粘液腺癌。
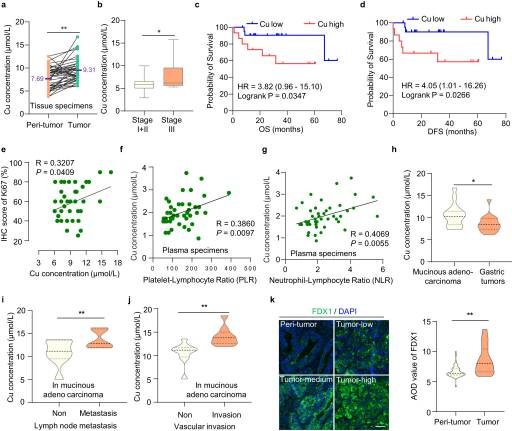
图1. 胃癌中铜含量过高,与肿瘤进展有关
2.METTL16是铜死亡的关键介质
有报道称,与良性胃病和健康对照患者相比,胃癌组的总m6A水平显著升高,并促进胃癌发生、生长、侵袭、上皮间充质转化(epithelial mesenchymal transformation, EMT)、转移甚至多药耐药的过程。然而,m6A修饰与铜突的关系仍不清楚。有趣的是,我们发现铜含量与GC组织中总m6A水平呈正相关(R = 0.5116)(图2A)。此外,铜显著上调GC细胞中m6 A修饰的总水平(图2B)。考虑到甲基转移酶样蛋白(METTL)家族在促进m6A修饰中起主导作用,我们分析了以1:1比例使用埃司氯莫尔/双硫拉姆和铜处理METTL敲低细胞的活力。结果表明,METTL16敲低导致对埃司克洛莫尔/双硫仑-铜处理产生耐药性(图2C-E)。此外,四硫钼酸盐(TTM)是一种铜体变形抑制剂,在对照细胞和METTL16敲低的细胞中抑制了铜体变形,表明METTL16参与了铜体变形(图2F)。观察到的结果与METTL16敲除克隆细胞系相似(图2G, H)。这些结果表明,METTL16在铜增生中起着至关重要的作用。
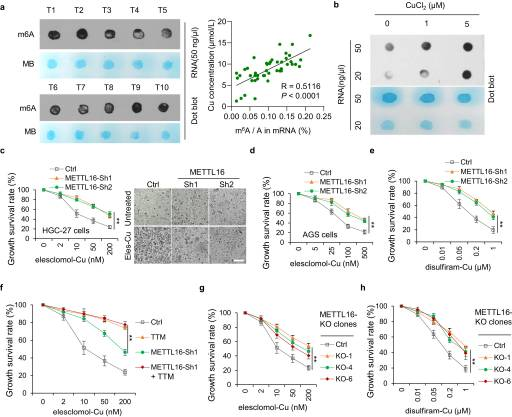
图2. METTL16是铜死亡的关键介质
3.METTL16通过靶向FDX1促进铜死亡
METTL16在许多转录中负责m6 A的沉积。因此,在稳定的稳敲对照和稳敲METTL16细胞系中进行甲基化RNA免疫沉淀(MeRIP)测序(图3A)。。使用HOMER软件预测每个样品中潜在的m6 A修饰,m6 A修饰主要映射到经典的GGAC基序(图3B)。GO富集分析显示,mettl16修饰基因富集了前15条通路,其中一些与线粒体通路相关(图3C)。此外,我们从铜质增生相关通路中发现了10个与METTL16稳敲组中m6A甲基化水平低以及mRNA水平下调相关的典型因子(图3D。qPCR分析显示,METTL16稳敲细胞中FDX1、MTF1、PDHB、ATP7A和DLD mRNA水平降低。与对照细胞相比,FDX1在METTL16-sh细胞中的mRNA水平下降最为显著(图3E)。此外,在METTL16- sh或-KO细胞中检测到FDX1蛋白水平显著降低(图3F, G)。考虑到FDX1是铜离子载体诱导细胞死亡的关键调节因子,我们推测METTL16诱导的铜细胞凋亡依赖于FDX1。接下来,我们评估METTL16是否通过影响FDX1的表达来调节铜死亡。鉴于蛋白质脂酰化是铜氧化的关键因素,我们使用硫辛酸特异性抗体作为DLAT脂酰化的测量,评估了METTL16敲低是否会影响蛋白质脂酰化。结果显示,METTL16敲低导致DLAT脂酰化降低,通过免疫印迹检测(图3H)。此外,在 METTL16-Sh 或 -KO 细胞中检测到 FDX1 蛋白水平显着降低 (3F,G)。考虑到FDX1是铜离子载体诱导细胞死亡的关键调节因子,我们推测METTL16通过FDX1诱导铜死亡。
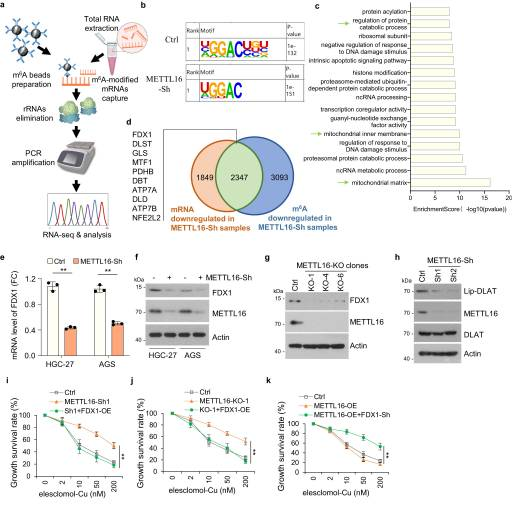
图3. METTL16通过靶向FDX1促进铜死亡
4.METTL16通过 FDX1 mRNA的m6A修饰促进 FDX1 的积累
为了进一步探讨METTL16如何调控m6 A修饰从而影响FDX1 mRNA水平,我们进行了基因特异性MeRIP-qPCR分析。我们发现,在METTL16敲除和敲除细胞中,FDX1 mRNA上的m6 A水平显著降低在转染mettl16过表达质粒的细胞中显著增加(图4A, B)。当使用放线菌素D (ActD)停止转录时,mettl16敲除细胞中FDX1 mRNA的衰减速度比对照细胞快(图4C)。。这些结果表明mettl16介导的甲基化导致FDX1 mRNA的稳定性。整合基因组学观察(IGV)分析FDX1富集的m6 A峰显示,与对照细胞(红色峰)相比,METTL16-sh细胞(绿色峰)中的m6 A水平降低,表明METTL16可能促进FDX1在CDS或3'UTR区域的m6 A修饰(图4D)。因此,我们将FDX1的部分CDS融合到pGL3荧光素酶报告基因下游,构建FDX1- wt荧光素酶报告基因。通过共转染FDX1荧光素酶报告基因和不同剂量的METTL16- oe质粒,我们发现METTL16显著提高了FDX1荧光素酶的活性(图4E)。
为了进一步探索它,我们使用SRAMP(基于序列的m6A修饰位点预测器)预测了FDX1 mRNA上可能的m6A修饰位点。综合图4e的预测结果和数据,我们以非常高的置信度确定了两个潜在的m6 A修饰位点:FDX1转录本的CDS区域602a位点和UTR区域820 A位点(图4F, G)。随后使用特异性引物进行MeRIP-qPCR分析,证明FDX1转录本上的602位点是mettl16介导的甲基化的直接底物(图4H)。此外,我们基于FDX1-WT荧光素酶报告基因,用胸腺嘧啶(T)取代m6A基序中的特异性腺苷(A),设计并构建了FDX1-602-Mut和FDX1-820-Mut荧光素酶报告基因(图4H)。双荧光素酶检测结果表明,METTL16不能促进602位点突变的FDX1-CDS/UTR报告基因构建体的荧光素酶活性(图4I)。对GEPIA数据库中METTL16和FDX1的表达分析表明,METTL16和FDX1在GC中呈正相关(图4j)。此外,免疫组织化学染色显示,在包含54对GC和邻近正常组织的组织芯片中,METTL16的表达与FDX1的表达呈正相关(图4K)。综上所述,这些结果证实了mettl16介导的FDX1 mRNA上的m6 A修饰对FDX1 mRNA的稳定性至关重要。
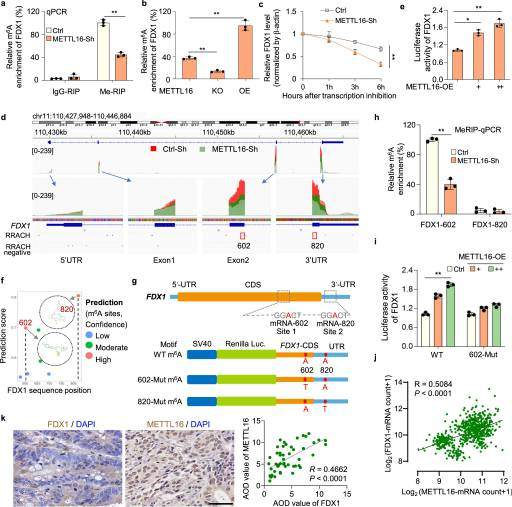
图4. METTL16通过 FDX1 mRNA的m6A修饰促进 FDX1 的积累
5.METTL16在铜胁迫下在 K229 位点乳酸化
接下来,我们评估了METTL16是否对GC中的铜胁迫有反应。铜处理后,METTL16 mRNA和蛋白水平不变,但FDX1蛋白水平显著升高(图5A, B)。为了验证铜胁迫下FDX1表达上调是否依赖于METTL16,我们测量了在存在或不存在铜的情况下,METTL16敲除或敲除细胞中FDX1蛋白的表达。结果显示,METTL16缺乏消除了铜诱导的FDX1上调(图5C, D)。由于METTL16 mRNA和蛋白水平在铜胁迫下保持不变,我们随后使用质谱法测量了METTL16 PTM的变化。分析显示,METTL16蛋白序列上有11个乳酸化位点和2个乙酰化位点(图5E)。乳酸处理后,6个乳酸化位点(红色)的PTM水平显著升高,提示这6个位点在肿瘤细胞中可能受到调控。然后我们证实了对铜胁迫的乳酸化或乙酰化反应的存在。结果显示,铜处理显著提高了METTL16的乳酸化水平,而不是乙酰化水平(图5F)。为了确定METTL16在铜相关代谢中的重要乳酸化位点,我们生成了这6个乳酸化位点的乳酸化缺陷突变体(K ~ R)和K410(作为代表位点,其乳酸化在乳酸胁迫下保持不变)。我们发现只有METTL16-K229R突变体在铜胁迫下显示乳酸化减少(图5G)。随后,通过LC-MS/MS分析得到METTL16-K229与其他乳酸化位点的b-y离子匹配图(图5H)。为了进一步证实铜胁迫下METTL16-K229的乳酸化,我们生成了一种METTL16乳酸-K229抗体,并利用乳酸-K229肽和表达METTL16- wt、-K229R或-K229E的细胞来验证其有效性和特异性(图5I, J)。铜和乳酸处理均诱导K229发生强烈的METTL16乳酸化(图5k, l),表明K229是铜相关代谢中必需的乳酸化位点。这些结果表明,铜胁迫下METTL16在K229位点发生乳酸化。在稳定的METTL16拯救(METTL16-WT、-K229R或-K229E)细胞系中,内源性METTL16被野生型(WT)、乳酸化缺陷(K229R)或乳酸化模拟(K229E)取代,进一步确保了FDX1蛋白水平。结果显示,mettl16敲除细胞中FDX1蛋白水平下降,在METTL16-WT或-K229E拯救细胞后恢复,但METTL16-K229R拯救细胞中没有恢复(图5M)。综上所述,铜胁迫诱导了METTL16- k229的乳酸化,并促进了METTL16的甲基转移酶活性。
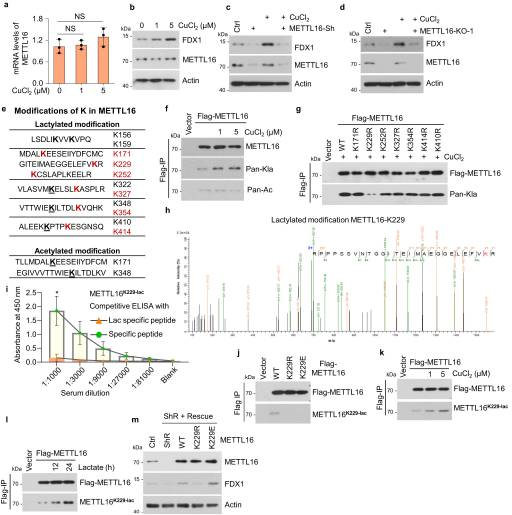
图5. METTL16在铜胁迫下在 K229 位点乳酸化
6.SIRT2 使 METTL16-K229 脱乳酸并抑制METTL16活性
SIRT2被鉴定为METTL16的结合蛋白,是一种典型的去乙酰化酶,分为主要的去乙酰化酶和乳酸转移酶组(图6A)。EIF3b也被鉴定为METTL16的结合蛋白,与以往的研究一致29。为了研究和验证METTL16和SIRT2之间的相互作用,在HGC-27细胞中检测到METTL16-SIRT2结合(图6B)。使用His-pull-down试验验证了METTL16-SIRT2的直接关联(图6C)。METTL16-K229的乳酸化被SIRT2过表达显著抑制(图6D),并被SIRT2敲低或用SIRT2抑制剂- AGK2处理促进(图6E)。
此外,铜诱导的METTL16-K229的乳酸化被SIRT2过表达抑制,而被SIRT2敲低或AGK2处理增加(图6F, G)。总的来说,这些结果表明SIRT2与K229位点的METTL16相互作用并使其去乙酰化。我们进一步探讨了SIRT2在稳定的METTL16拯救(METTL16- wt, -K229R或-K229E)细胞系中对METTL16的调控作用。SIRT2过表达抑制了METTL16- wt细胞中的FDX1 m6 A水平,但在METTL16- k229r或-K229E细胞中没有,这表明SIRT2通过METTL16- k229的乳酸化抑制了METTL16的活性(图6H, I)。此外,SIRT2过表达抑制了FDX1蛋白水平(图6J),而SIRT2敲低和AGK2处理促进了FDX1蛋白水平(图6K)。免疫荧光染色(IF)结果显示,GC组织芯片中SIRT2蛋白水平与FDX1蛋白水平呈负相关(图6l)。
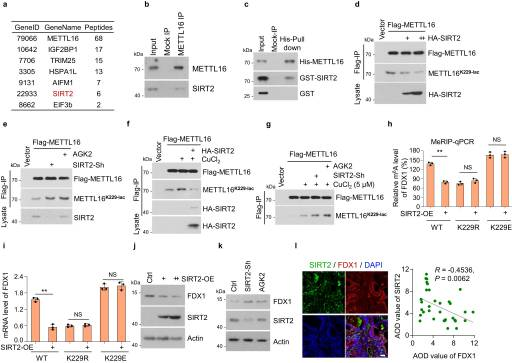
图6. SIRT2 使 METTL16-K229 脱乳酸并抑制METTL16活性
7.SIRT2-METTL16-FDX1-铜死亡轴支持一种有前途的治疗策略
FDX1是铜质增生的重要介质,铜质增生是一种潜在的癌症治疗方法53。我们发现,在sirt2抑制或铜诱导的METTL16-K229乳酸化后,FDX1蛋白水平升高。接下来,我们研究了METTL16-K229乳酸化对铜生长的影响。结果显示,铜胁迫下METTL16敲低介导的DLAT脂酰化降低在METTL16- wt /-K229E细胞中得以恢复,但在METTL16- k229r细胞中没有恢复(图7A)。此外,在铜存在的情况下,elesclool处理在METTL16-WT或-K229E细胞中诱导铜变性,但在METTL16-K229R细胞中没有(图7B)。此外,SIRT2过表达逆转了铜胁迫下METTL16-WT诱导的DLAT脂酰化的增加,但没有逆转METT16-K229E诱导的DLAT脂酰化,这表明SIRT2通过使METTL16K229去乙酰化来抑制cupropsis(图7C)。与此一致的是,AGK2处理促进了METTL16-WT细胞中的DLAT脂酰化,而在铜胁迫下的METTL16-K229R细胞中则没有(图7D)。此外,在埃司克洛莫尔和铜存在的情况下,AGK2治疗促进铜增生(图7E)。我们进一步评估了METTL16乳酸化是否能增强接受埃司洛莫尔治疗的小鼠异种移植肿瘤的治疗反应。将METTL16-WT、-K229R、-K229E细胞皮下注射到裸鼠左右后腿上方。METTL16-WT组和-K229E组小鼠肿瘤生长明显受到抑制,而METTL16-K229R组则没有,通过肿瘤体积和重量的减少可以观察到(图7F-H)。肿瘤切片FDX1染色在METTL16-WT和-K229E组中高于METTL16-K229R组(图7I)。这些结果表明,K229位点METTL16的乳酸化状态在铜质增生的决定中起着至关重要的作用。此外,在异种肿瘤移植中使用埃司克洛莫尔和AGK2联合治疗。肿瘤切片分析显示,AGK2在体内对埃司氯莫尔治疗增敏,通过肿瘤体积和重量的减少可以观察到(图7J - L)。这些结果表明,埃雷斯洛莫尔和AGK2联合治疗胃癌,特别是恶性肿瘤-粘液腺癌(高铜含量)是有希望的治疗方法。
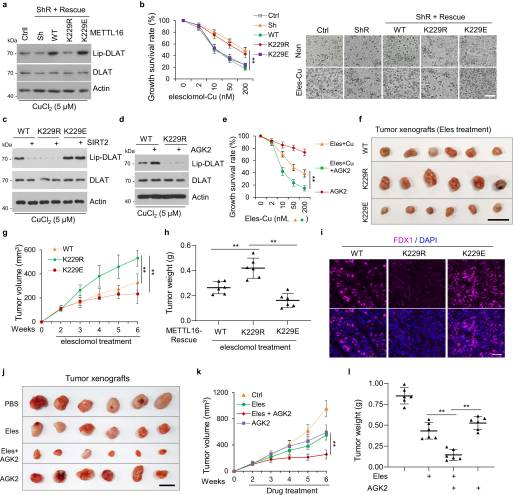
图7.SIRT2-METTL16-FDX1-铜死亡轴支持一种有前途的治疗策略
结论:
在这里,我们证明了研究团队发现铜离子促进了胃癌的整体m6A修饰,其中METTL16介导的FDX1 mRNA - m6A在促进铜死亡方面起到关键作用。具体而言,铜离子通过调控乳酰化酶和去乳酰化酶SIRT2 调控METTL16-K229乳酰化,从而调控其活性。乳酰化的METTL16引起FDX1-602位点发生甲基化修饰,促进FDX1表达,导致铜死亡发生。SIRT2是一种典型的去乙酰化酶,其抑制剂AGK2在该途径中可以显著促进METTL16的乳酰化及FDX1的m6A修饰,促进铜死亡。因此,AGK2联合铜死亡诱导剂伊利司莫(Eles),可以促进恶性胃癌的治疗。
实验方法:
WB、q-RTPCR、异种移植瘤实验、稳定细胞系构建、m6斑点印迹测定、荧光素酶报告基因检测、His-pull-down测定、IP、免疫荧光、MeRIP-qPCR、甲基化RNA免疫沉淀测序、RIP、体外乳酸化测定
参考文献:
Sun L, Zhang Y, Yang B, et al. Lactylation of METTL16 promotes cuproptosis via m6A-modification on FDX1 mRNA in gastric cancer. Nat Commun. 2023;14(1):6523. Published 2023 Oct 20.