






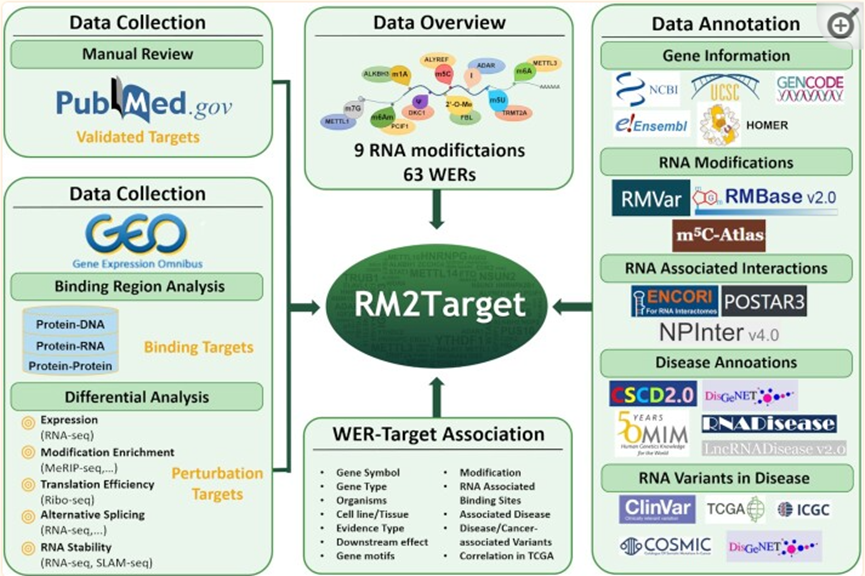
RM2Target:RNA修饰数据库
RNA修饰在调节RNA代谢中发挥着关键作用,如基因转录、RNA稳定性、RNA剪接、核定位和翻译。RNA修饰是由三类不同的蛋白质动态介导的:写入者、擦除者和读取器(WER)。写入者可以调节RNA修饰的沉积;例如,METTL3和NSUN2可以分别将N6-甲基腺苷(m6A)和5-甲基胞嘧啶(m5C)写入其靶基因。擦除者去除靶基因的RNA修饰;例如,FTO是m6A的擦除器,而ALKBH3是N1甲基腺苷(m1A)的擦除器。读取器通过识别靶基因中的RNA修饰位点来发挥其功能;例如YTHDF1/2/3是m6A(6)的读取器并且ALYREF是m5C的读取器。WER的失调已被证明与各种疾病有关,包括癌症、心血管疾病和神经系统疾病。相同的WER在不同的条件下可能具有不同的功能。例如,METTL3在大多数癌症类型中起致癌作用,但据报道,它在某些癌症类型中也具有肿瘤抑制功能,如肾癌。这主要是因为WER的扰动可能在不同的条件下选择性地影响不同的靶基因集。总之,识别WER-靶标关联对于研究RNA修饰在各种生理和病理条件下的功能和调节机制尤为重要。然而,目前还没有公共存储库来承载不同RNA修饰的WER-靶标关联。
得益于实验和高通量测序技术的发展,可以为探索WER与靶基因之间的关系提供更多的证据。例如,免疫沉淀和某些下一代测序技术,如RIP-seq和CLIP-seq,可以直接有效地阐明WER蛋白与靶RNA之间的结合关系。微扰技术,如小干扰RNA(siRNA)和CRISPR/Cas9系统以及高通量测序,可用于系统评估WERs对特定基因RNA代谢的微扰效应。在过去的两年里,与RNA修饰相关的研究激增。随着更多WER的发现,靶基因的数量也有所增加,因此需要更新m6A2Target。因此研究团队升级了数据库并取名为RM2Target (http://rm2target.canceromics.org/)。来自两种生物体和九种类型的RNA修饰的63个WER的靶标关联,包括N6甲基腺苷(m6A)、N6,2'-O-二甲基腺苷(m6Am)、N1甲基腺苷(m1A)、假尿苷(ψ)、5-甲基尿苷(m5U)、5-甲基胞嘧啶(m5C)、7-甲基鸟苷(m7G)、2'-O-甲基化和A-to-I RNA编辑。RM2Target含有包含1619653个WER,为了让用户进一步研究WER和靶基因的功能,并研究RNA修饰与疾病之间的关系,RM2Target提供了基本的基因信息和丰富的注释,包括RNA修饰、RNA-RNA/RNA-蛋白质相互作用、TCGA癌症类型中WER及其靶基因之间的表达相关性,以及RNA与疾病的关联。
三种证据类型的WER-靶标关联存储在RM2Target中:(i)“已验证”:分别使用蛋白质印迹、RT-qPCR、RNA稳定性测定、RIP测定、荧光素酶报告基因测定等方法,通过体内或体外实验验证人类和小鼠的1530和584个WER-靶点关联。(ii)“结合”:通过对人类和小鼠的高通量分析,分别预测了461746和61395 WER-靶标与结合证据的关联。其中,人类有451016个WER-靶标与蛋白质-RNA相互作用的关联,小鼠有55004个。在人类中总共记录了3100个WER-靶标与蛋白质-蛋白质相互作用的关联。(iii)“扰动”:分别从人类和小鼠的WER扰动中推断出646 539和447 859 WER-靶标关联。先前的研究表明,RNA修饰在某些基本的生物过程中发挥着至关重要的作用,如mRNA的稳定性、剪接和翻译。因此,RM2Target在五个水平上进行了差异分析,即表达、修饰、翻译、稳定性和选择性剪接。对于人类,有339604个WER-靶标与RNA水平变化相关,60176个与修饰水平变化有关,117866个与翻译效率变化有关,27700个与mRNA稳定性变化有关,101193个与差异选择性剪接事件有关。
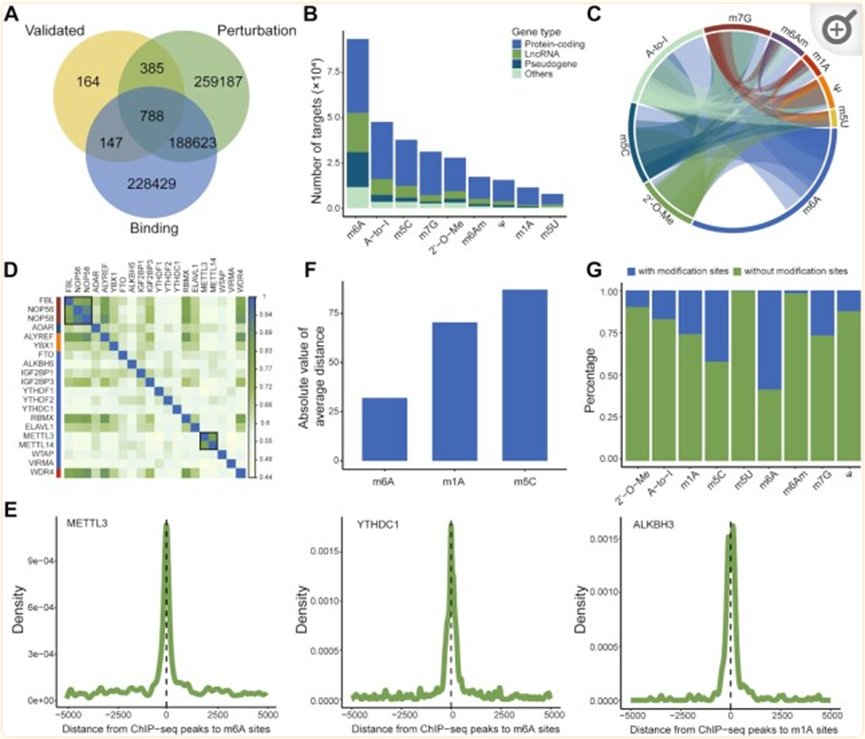
RM2Target揭示WER与靶基因之间的复杂调控网络
不同WER之间的串扰同一RNA修饰的写入者、擦除者和读取器经常合作控制细胞表型。越来越多的证据表明,不同的RNA修饰之间存在频繁的串扰。例如,NSUN2介导的m5C修饰可以促进METTL3介导的m6A修饰,反之亦然。m6A修饰可能通过阻断ADAR和靶基因的结合来抑制A-to-I编辑水平。我们系统地分析了在RM2Target中收集的数据,以探索不同RNA修饰的不同WER之间的串扰。观察到来自不同修饰的WER的靶基因之间的密集交联。正如预期的那样,m6A写入者METTL3和METTL14的靶基因具有高度重叠,与METTL3和METTL14通常作为异二聚体复合物起作用的事实一致。此外,FBL、NOP56和NOP58,2′-O-甲基化的作者,被证明是盒小核仁核糖核蛋白复合物(snoRNPs)的核心蛋白,也共享相似的靶基因集。
RNA修饰和基因转录之间的串扰RNA修饰的沉积通常被认为是一个共转录过程。RNA修饰也可以调节基因转录。通过对WER的ChIP-seq数据和RNA修饰位点的峰位点进行综合分析,我们发现m6A-WER的ChIP-seq峰位点,如METTL3、YTHDC1,靠近相应的m6A位点,m1A-WER峰位点,例如ALKBH3,接近相应的m1A位点,表明这些RNA修饰和基因转录之间存在强烈的串扰。我们计算了WER的ChIP-seq峰中心和最近的RNA修饰位点之间的平均距离,并认为该平均距离表明了RNA修饰和基因转录之间的串扰。因此,我们观察到m6A修饰主要涉及串扰,m5C最少涉及串扰。
与RNA修饰无关的WER-靶标关联最近的研究表明,一些已知的RNA修饰的WER具有与修饰无关的功能。例如,m6A写入器METTL3可以在没有m6A修饰的情况下促进靶基因PAPBC1的翻译。m6A写入物METTL16发挥m6A非依赖性功能以促进翻译和肿瘤发生。我们使用RM2Target数据系统地研究了RNA修饰依赖性和非依赖性WER-靶标关联。我们发现,只有30%的靶基因具有来自公共数据库(如RMbase和RMVar)的相应RNA修饰位点,这表明WERs可能具有RNA修饰无关的功能。然而,目前检测RNA修饰的方法可能由于各种原因错过了许多RNA修饰位点,这一可能性不容忽视。
