






肠道微生物代谢物通过抑制铁死亡促进结直肠癌的发展
肠道菌群在人体健康中起着关键作用。新出现的证据表明,肠道微生物通过产生致癌代谢物参与肿瘤发生的进展。然而,潜在的分子机制在很大程度上是未知的。在本研究中,作者发现从厌氧菌胃链球菌中提取的色氨酸代谢物,反式-3-吲哚丙烯酸(IDA),促进结直肠癌的发生。在机制上,IDA作为芳烃受体(AHR)的内源性配体,转录上调ALDH1A3(醛脱氢酶1家族成员A3)的表达,ALDH1A3利用视网膜作为底物产生NADH, NADH是铁氧化抑制蛋白1(FSP1)介导的还原性辅酶Q10合成所必需的。在体外和体内,AHR或ALDH1A3的缺失在很大程度上消除了IDA促进的肿瘤发展。有趣的是,厌氧菌在结直肠癌(CRC)患者中显著富集。在异种移植模型和ApcMin/+小鼠中,IDA治疗或厌氧消化链球菌的植入均可促进结直肠癌的进展。总之,作者的研究结果表明,靶向IDA-AHR-ALDH1A3轴应该有望用于铁衰相关的CRC治疗。本文章于2024年1月发表在《nature cell biology》,IF:21.3。
机制图:
主要研究结果:
1. IDA作为铁死亡抑制因子的鉴定
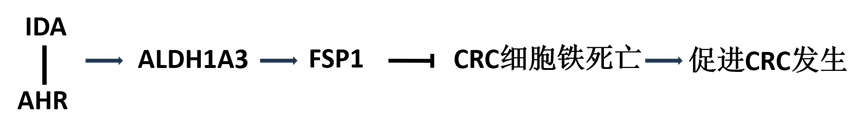
第一部分,为了测试肠道微生物代谢物是否会影响癌细胞铁死亡敏感性,作者在786-O人肾腺癌细胞中建立了包含350种内源性人宿主和肠道微生物代谢物的筛选库,用于检测铁死亡(图1a)。筛选数据确定了肠道微生物群色氨酸代谢产生的IDA,它显著抑制RSL3诱导的铁死亡(图1b)。为了验证这一发现,利用两种典型的对铁死亡敏感的癌细胞系786-O和HT1080人纤维肉瘤细胞来测试IDA的作用,发现IDA处理的细胞对铁死亡具有强大的抗性(图1c,d)。作者还检测了IDA培养HT1080细胞的脂质过氧化水平,发现IDA显著降低了脂质过氧化(图1e),表明IDA是一种有效的铁死亡抑制因子。
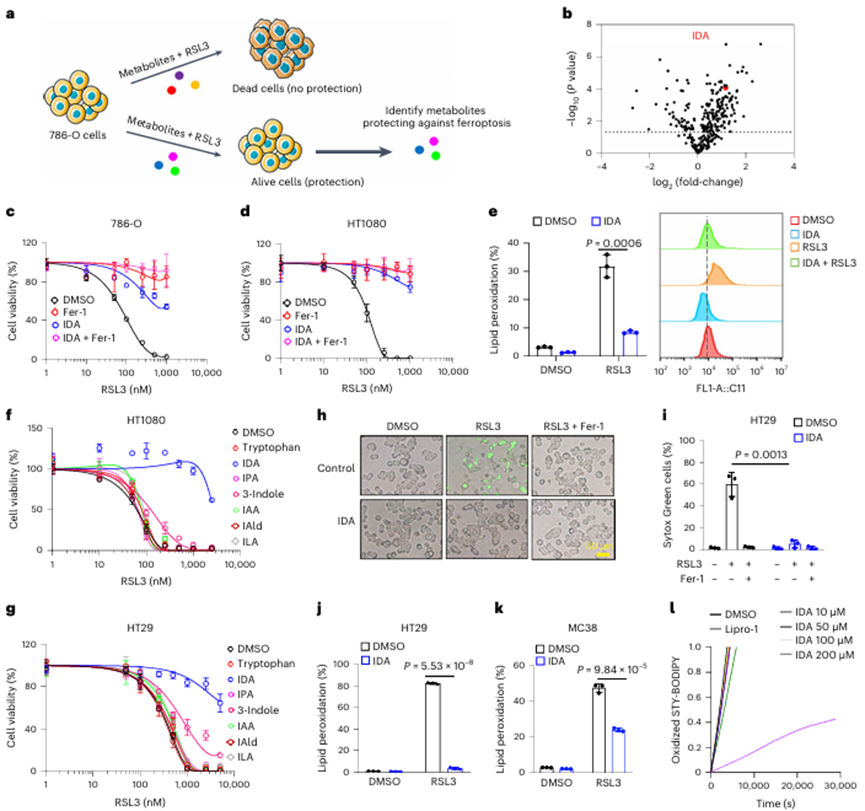
图1 IDA是一种有效的铁死亡抑制因子
肠道微生物群利用色氨酸合成多种衍生物,因此作者研究了其他肠道微生物来源的色氨酸代谢物是否可以调节铁死亡。发现,补充10 μM的IDA可以有效地消除HT1080细胞中的铁死亡,而低剂量(10 - 100 μM)的其他色氨酸衍生物则没有表现出很强的保护作用(图1f)。此外,补充IDA可以极大地消除HT29人结肠癌细胞中的铁死亡(图1g)。由于IDA在抑制铁死亡方面更有效,因此作者将注意力集中在IDA介导的铁死亡抑制上。在HT29细胞和MC38小鼠结肠癌细胞中,IDA处理显著抑制了高水平的铁死亡和脂质过氧化(图1h-k)。为了研究IDA是否作为自由基捕获抗氧化剂(RTAs)来抑制铁死亡,作者预测了所有色氨酸代谢物的亲脂性,表明IDA不能直接减少氧化脂质(图1l),提示IDA可能通过细胞信号通路介导铁死亡。
2. IDA促进结直肠肿瘤的发展
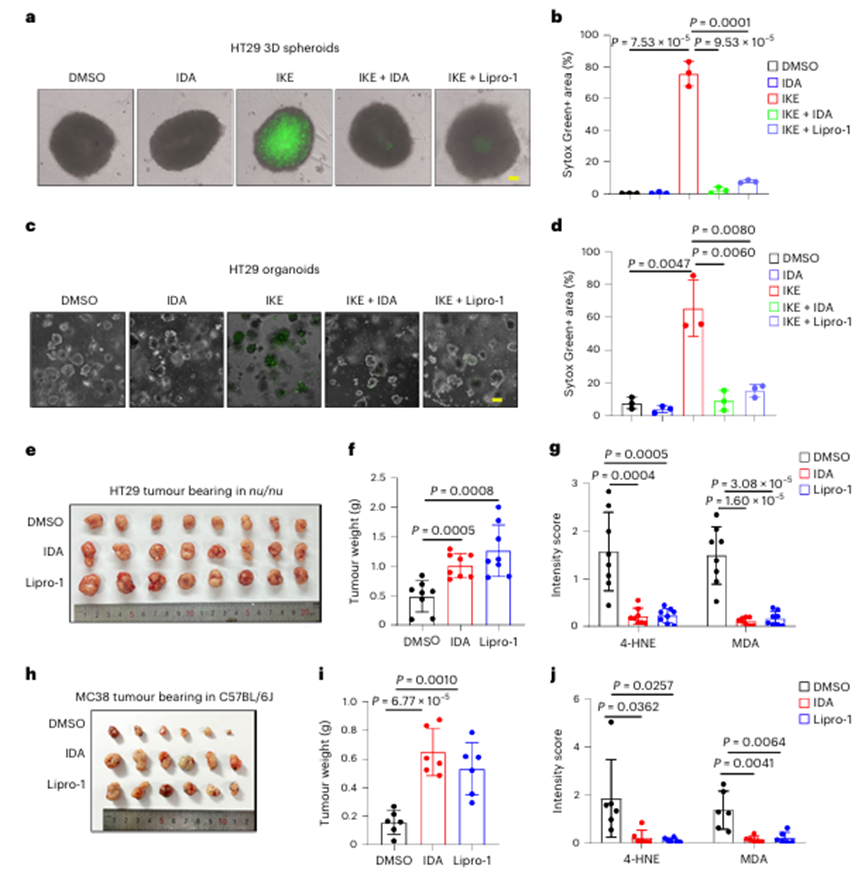
图2 IDA促进结直肠肿瘤的发展
由于体外二维细胞培养缺乏许多癌症的特征,如缺氧、细胞间接触改变和重新连接代谢,作者将HT29和MC38细胞培养成三维(3D)肿瘤球体,以更好地模拟体内环境。如图2a、b和扩展数据图2f所示,RSL3-或咪唑酮erastin (IKE)-诱导的HT29肿瘤球体铁死亡在很大程度上被IDA或脂前列腺素-1 (Lipro-1)所抑制。同样,在HT29类器官(图2c、d)中也观察到类似的结果。
为了确定IDA是否对肿瘤进展具有保护作用,作者进行了HT29异种移植,发现IDA显著地消除了脂质过氧化并促进了肿瘤发展(图2e-g)。与这一发现一致,IDA对C57BL/6J小鼠的铁死亡具有很强的保护作用,并促进MC38异种移植物的进展(图2h-j)。这些结果表明体内低浓度的IDA足以通过抑制铁死亡来促进肿瘤进展。
3. AHR是IDA介导的铁死亡抑制所必需的
先前的研究表明吲哚代谢物是AHR的幼稚配体。为了确定AHR是否参与IDA调控的铁死亡抑制,作者使用了AHR的拮抗剂BAY-218和StemRe genin 1 (SR1),发现AHR拮抗剂显著地消除了IDA调节的铁死亡抑制(图3a)。接下来,作者沉默了HT29和HT1080细胞中的基因表达(图3b)。IDA在表达 (sh)-AHR的细胞中未能抑制铁死亡(图3c)。此外,作者通过 (CRISPR)-Cas9技术敲除AHR(图3d)。由于缺乏AHR表达,IDA未能缓解铁死亡(图3e、f)。
此外,通过AHR的异位表达,IDA介导的AHR null HT29细胞中的铁死亡抑制得以恢复(图3g-j)。为了研究AHR是否有助于体内IDA介导的肿瘤发展,作者进行了野生型(WT)和AHR KO HT29异种移植实验,并证实IDA在AHR−/−细胞中的保护作用基本被消除(图3k-n)。
4. FSP1是IDA抗铁死亡作用所必需的
FSP1利用NAD(P)H催化还原性辅酶Q10的产生,可保护细胞免于铁死亡。因此,作者在HT29和HT1080细胞中生成了FSP1−/−克隆,以验证FSP1是否有助于IDA调节的铁死亡抗性。如图4a,b所示,HT29细胞中FSP1的KO在很大程度上取消了IDA-AHR轴的保护作用,而WT HT29细胞对IDA介导的铁死亡抑制有明显反应。同样,HT1080细胞中FSP1的缺乏完全阻断了IDA的抗铁死亡作用,类似于HT29 FSP1 sgRNA细胞(图4c-f)。相反,FSP1的异位表达挽救了这种表型(图4g,h)。此外,在HT29 WT细胞中,IDA的补充促进了肿瘤的发展,而在FSP1 KO细胞中,IDA的作用在很大程度上被取消(图4i,j),这表明IDA-AHR轴介导的铁死亡抑制依赖于FSP1。
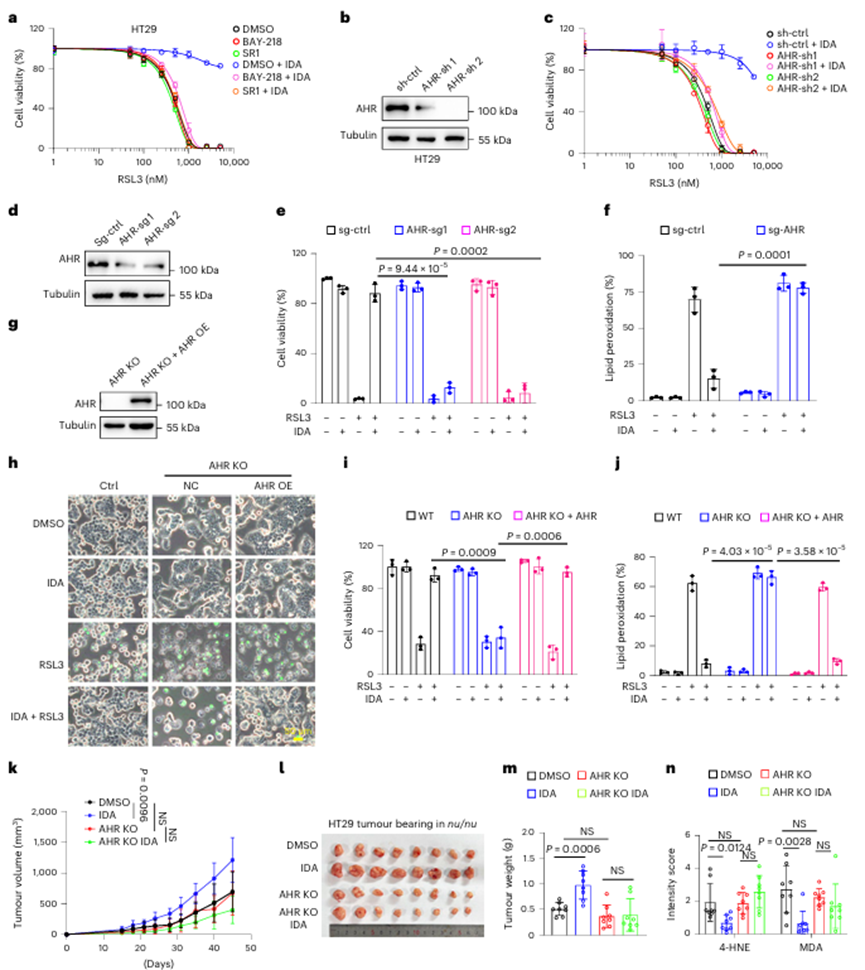
图3 IDA介导的铁死亡抑制需要AHR
5. IDA通过AHR-ALDH1A3-FSP1-CoQ10轴抑制铁死亡
上述发现提出了FSP1如何参与IDA-AHR介导的铁死亡抑制的问题。为了阐明这个问题,作者对IDA处理的HT29细胞进行了RNA测序(RNA-seq)。在IDA可能介导的基因中,ALDH1A3是上调最多的(图5a)。逆转录酶定量PCR (RT-qPCR)和western blotting分析证实,在IDA处理下,ALDH1A3的信使RNA和蛋白水平显著升高(图5b、c),这表明ALDH1A3可能是AHR的直接下游靶点。因此,作者接下来寻找ALDH1A3启动子周围潜在的AHR结合位点。通过与公认的AHR结合基序(GCGTG)比较,作者发现了三个潜在的结合区(R1-R3),其中R3包含一个相同的序列(图5d)。在接受或未接受IDA治疗的HT29中进行ChIP检测。有趣的是,在IDA处理下,内源性AHR在ALDH1A3转录起始位点(TSS)上游约100 bp处表现出与R3的强结合亲和力 (图5e),这表明在AHR被IDA激活的情况下,AHR在ALDH1A3启动子上募集。
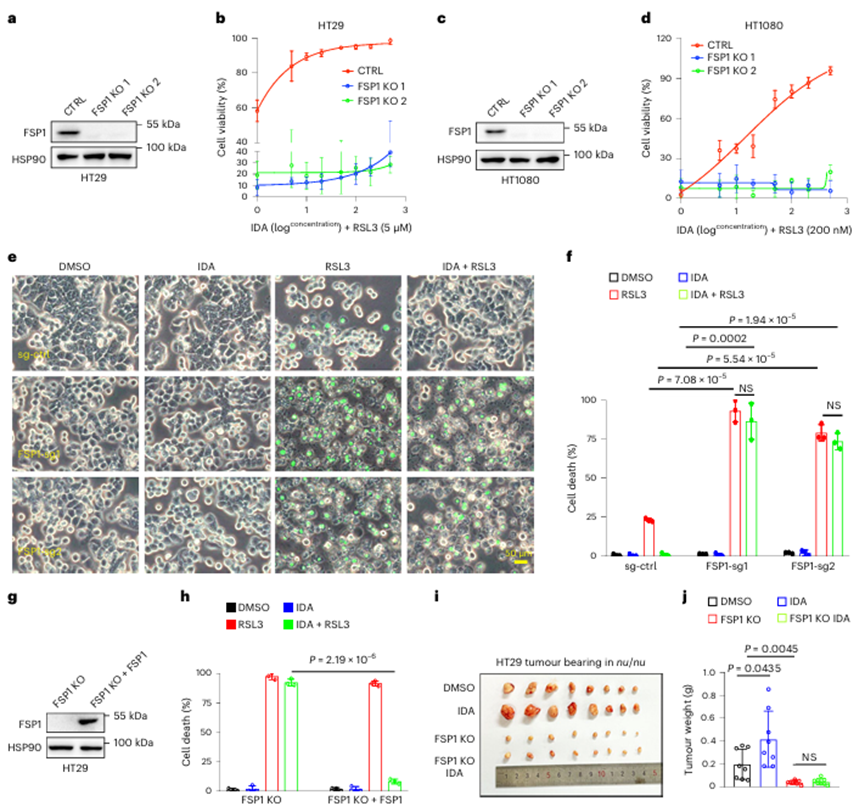
图4 IDA的抗铁死亡效应需要FSP1
为了检验ALDH1A3在铁死亡中的作用,作者阻断了HT29细胞中ALDH1A3的表达,并检测了铁死亡细胞的死亡水平。如图5f,g所示,ALDH1A3的缺失明显使细胞对脂质过氧化诱导的铁死亡敏感。此外,ALDH1A3表达降低会损害IDA抑制铁死亡的能力(图5h)。ALDH1A3特异性地利用全反式视黄醛作为底物产生NADH,视黄醛或维生素a调节的铁死亡保护依赖于ALDH1A3(图5i),表明视黄醛通过ALDH1A3的酶活性使细胞抵抗铁死亡。作者还使用ALDH1酶抑制剂双硫仑来测试ALDH1A3的作用,双硫仑诱导高水平的铁死亡细胞死亡,并减弱了IDA和ALDH1A3介导的铁死亡抑制作用(图5j)。

图5 IDA通过AHR-ALDH1A3-FSP1-CoQ10轴抑制铁死亡
为了研究ALDH1A3在肿瘤发展中的作用,作者测试了ALDH1A3的缺失或药理抑制是否会影响肿瘤细胞的生长。结果显示,ALDH1A3的缺失抑制了肿瘤的进展,阻断了IDA促进的肿瘤生长(图5k, l)。综上所述,作者的数据支持IDA-AHR通路介导的铁死亡抑制依赖于ALDH1A3。
为了进一步验证ALDH1A3产生的NADH是否参与FSP1介导的还原性辅酶Q10合成,作者首先研究了ALDH1A3产生NADH的酶活性。体外实验表明,纯化的ALDH1A3可以通过视黄醛产生NADH(图5m,n)。此外,ALDH1A3缺失显著降低了NADH水平,而在ALDH1A3 KO细胞中补充NADH可恢复铁死亡(图5o,p)。此外,在ALDH1A3−/−和FSP1−/−细胞中检测到较低比例的还原为氧化的CoQ10(图5q)。综上所述,这些数据表明ALDH1A3对于IDA-AHR-FSP1-COQ10介导的铁死亡抑制至关重要。
6. 厌氧消化链球菌是IDA生物合成的主要因素
为了研究IDA在结直肠癌发展中的来源,作者从健康人和结直肠癌患者的粪便中分离出微生物物种,并分别使用16S核糖体RNA基因测序和质谱(MS)进行代谢组学分析,对微生物群的位置进行了大规模表征。发现厌氧消化链球菌在CRC患者中显著富集(图6a)。MS代谢物分析显示,产生了可检测的IDA含量(图6b)。此外,CRC患者粪便中的IDA含量远高于健康人(图6c),厌氧消化链球菌的丰度与粪便中的IDA水平呈正相关(图6d)。
作者在厌氧条件下培养厌氧消化链球菌,并使用液相色谱-质谱技术检测上清液中各种色氨酸衍生物的水平。事实上,厌氧菌有效地利用色氨酸产生高水平的IDA和IPA(图6e,f)。作者观察到12 h时IDA的浓度达到15 μM左右(图6e)。此外,厌氧菌上清液处理后,HT29细胞中ALDH1A3和CYP1A2水平显著升高,而大肠杆菌上清液对HT29细胞无影响(图6g,h)。此外,在添加厌氧菌上清液的细胞中,铁死亡明显受到抑制(图6i)。AHR的缺失使其失效,而AHR的过表达恢复了其保护作用(图6j)。
先前的研究表明,厌氧消化链球菌特异性粘附在结直肠癌粘膜上,并且厌氧消化链球菌与结直肠癌细胞的直接相互作用使得产生足够浓度的IDA来抑制结直肠癌铁死亡成为可能。为了进一步证明体内IDA的生理浓度是否防御铁死亡并促进肿瘤进展,作者首先在体内进行了厌氧卟啉单胞菌体内注射,并检测了肿瘤中IDA的含量。IDA在肿瘤中的浓度(约5μM)有效地降低了CRC铁死亡并促进了肿瘤生长(图7a–d)。此外,IDA的生理浓度足以消除体内脂质过氧化和铁死亡(图7a-d)。综上所述,这些数据表明,生理浓度的IDA在体外和体内都能有效抑制铁死亡。
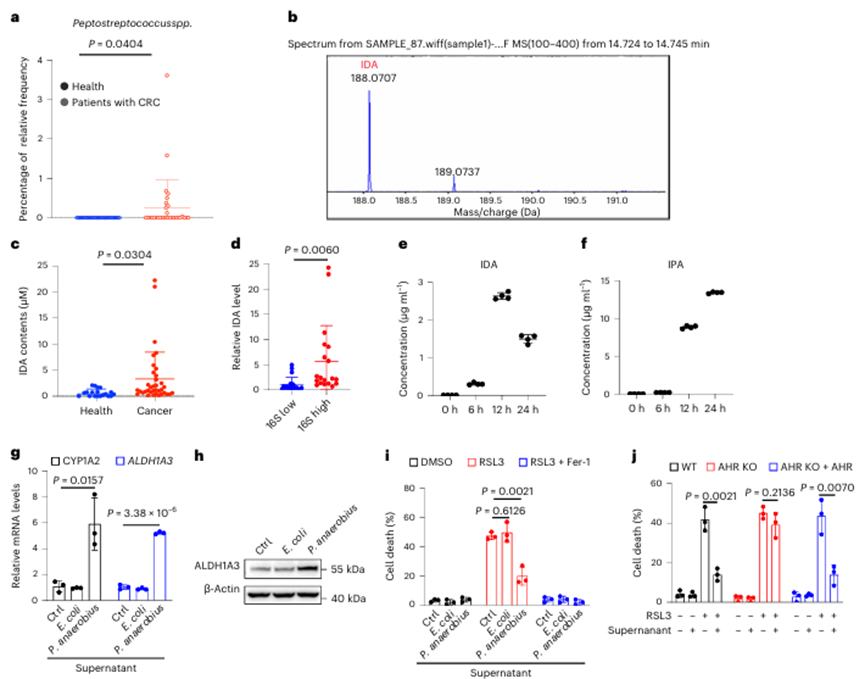
图6 厌氧菌是IDA生物合成的主要因子
7. 厌氧消化链球菌或IDA促进原位结直肠癌进展
为了更好地模拟体内肠道微生物群驱动的肿瘤发生,作者采用Apc突变(ApcMin/+)驱动的CRC模型来进一步研究。如图7e所示,厌氧消化链球菌在结肠炎症中表现出有效的定植(图7f)。此外,厌氧消化链球菌处理的ApcMin/+小鼠具有更高的肿瘤多样性(图7g-j),表明厌氧消化链球菌加速了ApcMin/+小鼠的结直肠发育。
作者进一步探讨厌氧消化链球菌介导的CRC发展是否依赖于IDA。为此,无菌小鼠被厌氧菌灌胃,这是更好地研究肠道微生物特征的有力工具。72h后,将这些小鼠安乐死,以检测血清色氨酸代谢物的丰度(图7k)。与体外实验结果一致,添加厌氧消化链球菌后,IDA和IPA显著增加(图7l,m),表明厌氧消化链球菌在体内负责IDA的生物合成。接下来,作者评估补充IDA是否会驱动ApcMin/+小鼠的结直肠肿瘤发生(图7n),经IDA处理的ApcMin/+小鼠显示出明显更高的肿瘤多样性(图7o)。进一步建立偶氮甲烷/葡聚糖硫酸钠(AOM/DSS)结肠炎相关癌症模型,发现IDA和厌氧p显著促进结肠炎诱导的CRC进展(图7p,q)。
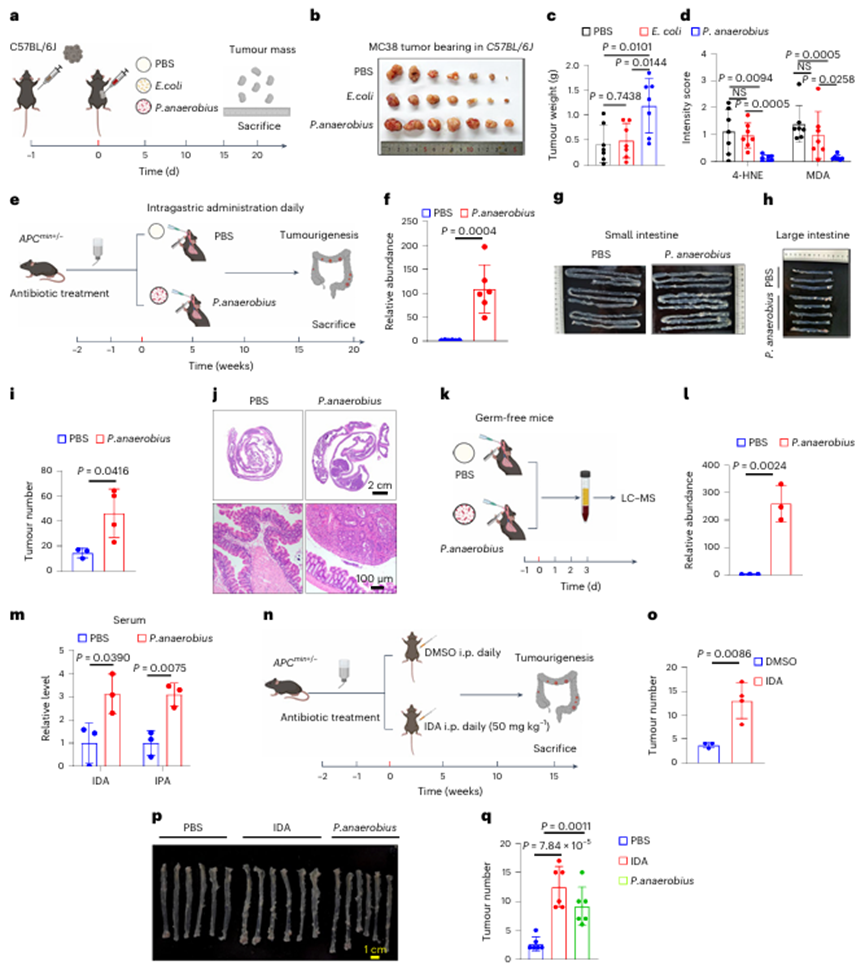
图7 厌氧消化链球菌或IDA促进原位CRC进展
接下来,作者分析了AHR在临床样本中的功能后果,发现AHR和ALDH1A3在结直肠癌中表达上调(图8a-c)。此外,AHR的高表达与CRC患者临床预后差呈正相关(图8d)。免疫组织化学(IHC)和western blotting分析一致显示,AHR和ALDH1A3在人类CRC样本中表达更高(图8e-h)。此外,4-羟基-2-壬烯醛(4-HNE)、3,4-亚甲基二氧安非他明(MDA)和8-羟基-2 ' -脱氧鸟苷(8-OHDG)的免疫组化染色显示,结直肠癌中脂质过氧化水平较低(图8i-k)。总之,作者的研究表明IDA-AHR-ALDH1A3 FSP1通路通过抑制铁死亡促进结直肠癌的发展。
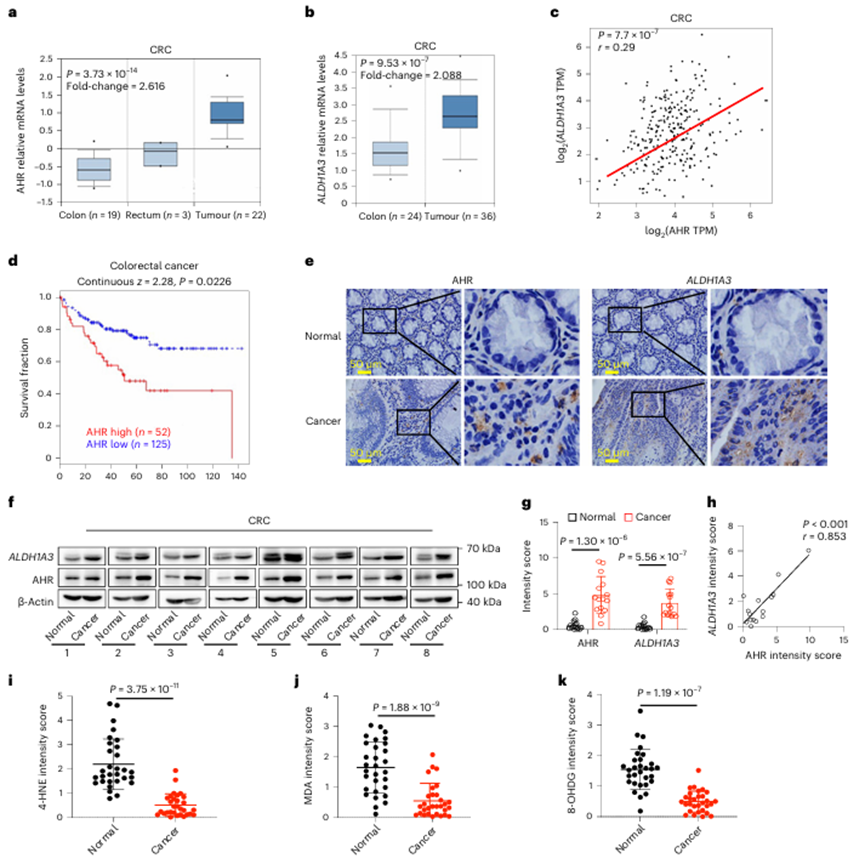
图8 AHR-ALDH1A3通路与CRC患者预后呈负相关
在本研究中,作者证明了IDA在CRC发展中的重要作用。具体来说,作者的工作表明,IDA作为AHR的内源性配体,转录上调ALDH1A3的表达,ALDH1A3产生的NADH用于FSP1介导的还原辅酶Q10的合成。CRC细胞中AHR或ALDH1A3的缺乏极大地抑制了IDA促进的肿瘤发展。作者的研究对于了解不同色氨酸代谢物在肿瘤进展中的作用具有重要意义。
实验方法:
细胞培养和稳转细胞系构建,细菌的分离、培养和鉴定,WB,细胞活力、细胞死亡及流式细胞术测定,DPPH检测,FENIX检测,3D肿瘤模型构建,类器官构建,质粒的构建和转染,定量PCR,ChIP检测,细胞质和细胞核分离,NADH水平检测,CoQ和CoQH2分析,LC–MS分析。
参考文献:
Cui W, Guo M, Liu D, Xiao P, Yang C, Huang H, Liang C, Yang Y, Fu X, Zhang Y, Liu J, Shi S, Cong J, Han Z, Xu Y, Du L, Yin C, Zhang Y, Sun J, Gu W, Chai R, Zhu S, Chu B. Gut microbial metabolite facilitates colorectal cancer development via ferroptosis inhibition. Nat Cell Biol. 2024 Jan;26(1):124-137. doi: 10.1038/s41556-023-01314-6. Epub 2024 Jan 2. PMID: 38168770.