






半凝集素-9在缺氧脑肿瘤微环境中介导小胶质细胞的功能
半乳糖凝集素-9是多种病理生理过程的多方面调节因子,以上下文依赖的方式发挥积极或消极的作用。在这里,作者阐明了半乳糖凝集素-9在脑肿瘤微环境中对髓系细胞的独特功能特性。半凝集素-9在肿瘤浸润细胞的小胶质细胞和巨噬细胞中高表达。在原代胶质细胞中,半乳糖凝集素-9的表达和分泌均受肿瘤的影响。对人类胶质母细胞瘤大量RNA测序数据集和来自小鼠胶质瘤模型的单细胞RNA测序数据集的分析揭示了半乳糖凝集素-9表达与胶质细胞活化之间的相关性。在脑肿瘤微环境中,半凝集素-9高的小胶质细胞亚群在功能上不同于半凝集素-9阴性/低的小胶质细胞亚群。高半乳糖凝集素-9的小胶质细胞表现出炎症激活和更高的细胞死亡率,而低半乳糖凝集素-9的小胶质细胞对脑肿瘤细胞表现出优越的吞噬能力。在小鼠胶质瘤模型中,阻断半乳糖凝集素-9抑制肿瘤生长并改变胶质细胞和T细胞的活性。这些发现表明骨髓细胞上的半乳糖凝集素-9是一种免疫调节剂,也是脑肿瘤的推定治疗靶点。该文章于2024年11月发表在《cancer research》,IF:14.2。
摘要图:

主要研究结果:
1. 半乳糖凝集素-9在颅内小鼠肿瘤模型的荷瘤同侧半球高表达
半乳糖凝集素-9似乎作为病理生理过程的多方面调节因子,以上下文依赖的方式发挥积极或消极的作用。为了更好地了解脑TME的免疫景观,作者使用颅内GL26小鼠胶质瘤模型研究了半乳糖凝集素-9的表达特征。WB显示,与对侧半球相比,雌性和雄性小鼠的同侧半球中半乳糖凝集素-9的表达明显更高(图1A)。在使用B35细胞的大鼠模型中观察到类似的结果(图1B和C)。免疫组织化学显示,半乳糖凝集素-9在同侧区域的表达明显高于对侧区域(图1D)。
接下来,作者使用表达EGFP的GL26小鼠研究了哪些细胞类型表达半乳糖凝集素-9。免疫组化显示,半乳糖凝集素-9在EGFP阳性的肿瘤细胞和肿瘤周围细胞上均有表达(图1E)。如图1F和G所示,半乳糖凝集素-9与CD3+ T细胞、IBA-1 +小胶质细胞和GFAP+星形胶质细胞共定位,在肿瘤边缘(即肿瘤病变与正常组织之间的界面)比肿瘤核心更明显。然而,作者没有在new +神经元中检测到半乳糖凝集素-9。为了验证这些发现,作者检测了GL26小鼠肿瘤浸润免疫细胞中半乳糖凝集素-9的水平。流式细胞术显示,与对侧肿瘤区域相比,同侧肿瘤区域的小胶质细胞(CD45midCD11b+)、巨噬细胞(CD45highCD11b+)和淋巴细胞(CD45highCD11b -)中的半凝集素-9水平显著增强(图1H)。这些结果表明半乳糖凝集素-9可能在脑TME内肿瘤周围免疫细胞中发挥特定作用。
2. 小胶质细胞半凝集素-9的表达在脑肿瘤中发生改变,并与炎症事件有关
小胶质细胞是至关重要的脑驻留前哨细胞,在第一道防线中起着至关重要的作用,脑肿瘤肿块通常由高达40%的髓系细胞组成,包括小胶质细胞。因此,作者主要关注小胶质细胞中的半凝集素-9,研究了小胶质细胞半凝集素-9的表达是否可以被脑肿瘤调节。半乳糖凝集素-9的转录物和蛋白水平在暴露于脑肿瘤CMs的原代胶质细胞中明显升高。随着时间的推移,在初级小胶质细胞中观察到这种增加(图2A)。FACS和免疫细胞化学分析进一步显示,与PACM相比,gl26cm处理的小胶质细胞细胞质和细胞表面半乳糖凝集素-9水平显著升高(图2B)。原代小胶质细胞与GL26-EGFP细胞共培养也导致小胶质细胞半乳糖凝集素-9的升高(图2C)。此外,间接共培养增强了半乳糖凝集素-9的表达和Ki-67+增殖小胶质细胞的比例。然后,作者监测原代胶质细胞分泌半乳糖凝集素-9,发现gl26cm处理的胶质细胞比PACM处理的胶质细胞分泌更多(图2D),表明肿瘤源性因子以旁分泌模式影响胶质半凝集素-9。
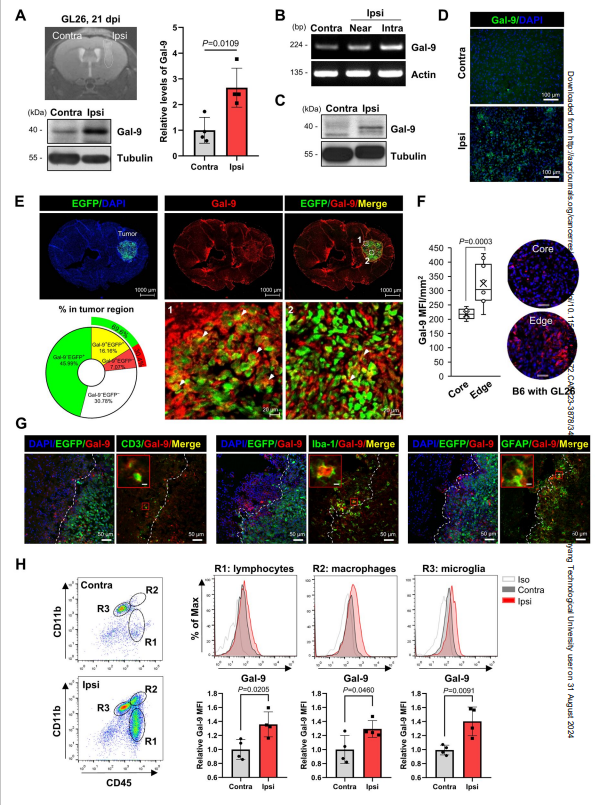
图1 半凝集素-9在颅内肿瘤模型的同侧半球表达升高
据报道,分泌的半乳糖凝集素-9在某些病理条件下作为一种损伤相关分子模式(DAMP)。为了评估半乳糖凝集素-9在脑TME中的功能特性,作者使用重组半乳糖凝集素-9(rmGal-9)检测了外源性半乳糖凝集素-9在免疫和炎症反应中的作用。用rmGal-9治疗小胶质细胞显著增加了半乳糖凝集素-9的转录物和蛋白水平,以及白细胞介素(IL)-1和IL-6等炎症细胞因子(图2E-G)。FACS分析显示rmGal-9升高炎症相关分子的水平,包括toll样受体(TLR)-2和程序性死亡配体1 (PD-L1,图2H)。
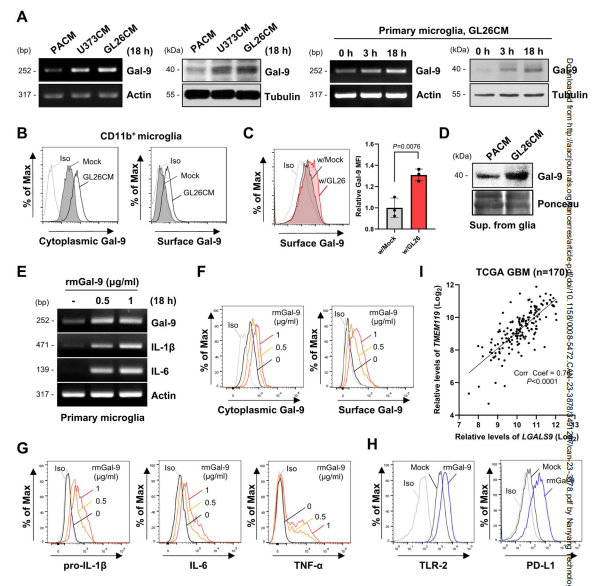
图2 小胶质细胞半凝集素-9在脑肿瘤中表达上调
这些发现促使作者研究脑TME中半乳糖凝集素-9表达与神经胶质活化之间的关系。作者分析了最初由Ochocka等人获得的小鼠GL261胶质瘤CD11b+细胞的scRNA-seq数据。与对照组相比,肿瘤组织中的小胶质细胞和胶质瘤相关巨噬细胞的Lgals9表达更高。使用MG评分,定义为小胶质细胞激活特征基因- TMEM119, P2ry12, Cx3cr1, Olfml3, Sparc和Gpr34,作者发现Lgals9水平与小胶质细胞簇中的小胶质细胞激活之间存在显著相关性。同样,TCGA-GBM分析显示Lgals9表达与TMEM119之间存在强正相关(图2I)。综上所述,这些结果表明神经胶质半乳糖凝集素-9的表达和分泌受到脑肿瘤的影响,半乳糖凝集素-9在脑TME内肿瘤暴露的神经胶质中诱导炎症反应。
3. 半凝集素-9高的小胶质细胞比半凝集素-9阴性/低亚群表达更高水平的炎症介质
鉴于上述发现,作者试图探索脑TME中半乳糖凝集素-9高表达和负/低表达的胶质细胞的功能特征。作者首先检查了半乳糖凝集素-9高和半乳糖凝集素-9阴性/低的小胶质细胞中代表性炎症相关分子的水平。与上述结果一致,在CD11b+半凝集素-9高的小胶质细胞中,TLR-2、TLR-4和PD-L1的水平明显高于CD11b+半凝集素-9阴性/低的小胶质细胞(图3A)。相反,半乳糖凝集素-9高和半乳糖凝集素-9阴性/低小胶质细胞亚群的TIM-3水平无显著差异。接下来,作者分析了gl26cm处理的TLR-2高表达小胶质细胞和PD-L1高表达小胶质细胞中半乳糖凝集素-9的水平。如图3B和C所示,与模拟处理的对照组相比,GL26CM显著增加了TLR-2高水平的小胶质细胞的比例,并且这些TLR-2高水平的细胞比TLR-2阴性/低水平的小胶质细胞表现出更高的半凝集素-9水平。同样,gl26cm诱导的高PD-L1小胶质细胞比暴露于GL26CM的低PD-L1亚群显示出更高的半乳糖凝集素-9水平(图3D和E)。这些发现进一步支持了半乳糖凝集素-9的表达与脑TME中小胶质细胞的炎症特性密切相关。
4. 半凝集素-9高的小胶质细胞比半凝集素-9阴性/低的小胶质细胞有更高的细胞死亡率
据报道,半乳糖凝集素-9可触发T细胞凋亡,而半乳糖凝集素-9相关的细胞凋亡与几种自身免疫性疾病有关。因此,作者询问半乳糖凝集素-9是否有助于脑TME中的小胶质细胞死亡。作者首先研究了分泌的半凝集素-9是否会影响神经胶质细胞的死亡。rmGal-9处理导致小胶质细胞增殖明显减少,Annexin-V + PI+凋亡细胞显著增加(图4A)。然后,作者探讨了rmGal-9是否与caspase-1激活介导的炎症相关细胞死亡有关。FAM-FLICA和FVS-780染色显示,rmGal-9显著升高活细胞中FLICA+活性caspase-1,显著增加小胶质细胞中FLICA+ FVS-780+炎性细胞死亡(图4B)。WB和免疫细胞化学染色进一步表明,与模拟处理的对照组相比,rmgal -9处理的胶质细胞表现出气皮素-d(GSDMD)的大幅上调,GSDMD是炎症性半胱天蛋白酶的生理底物和热相关调节因子,其活性裂解形式(图4C)。这些结果表明,细胞外半乳糖凝集素-9可能引发细胞凋亡和炎症相关的胶质细胞死亡。
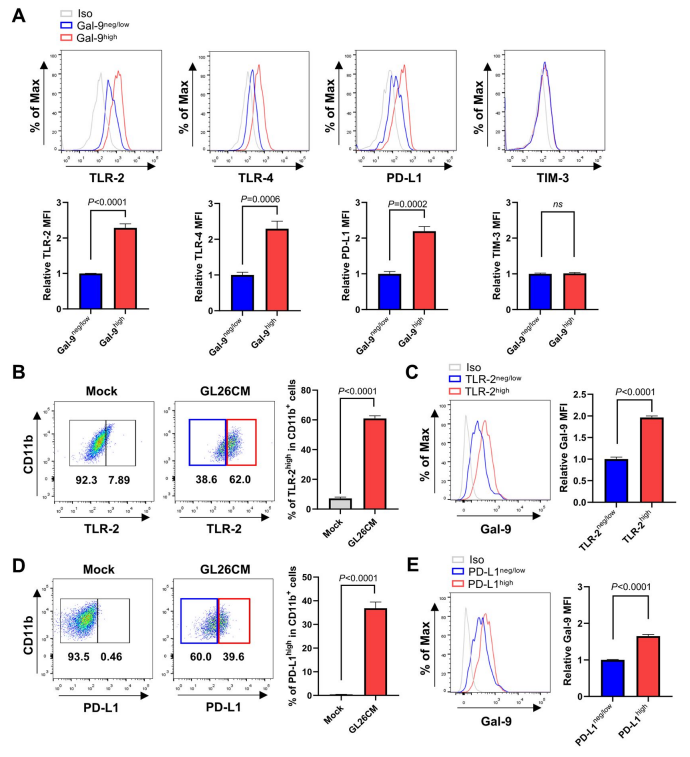
图3 半凝集素-9影响神经胶质细胞的炎症免疫反应
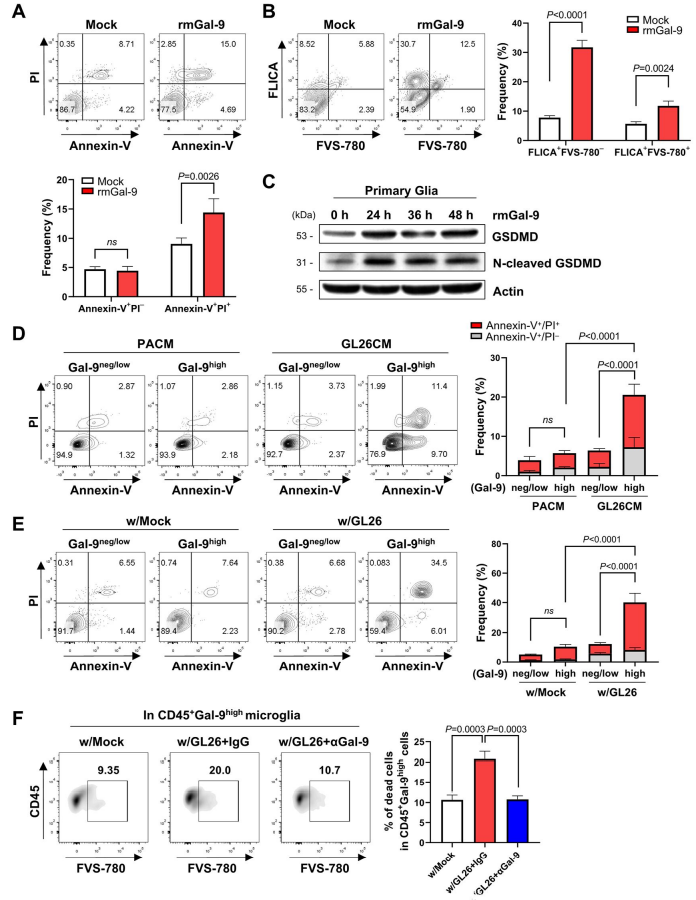
图4 小胶质半凝集素-9有助于脑TME中的细胞死亡
接下来,作者比较了半凝集素-9高和半凝集素-9阴性/低小胶质细胞亚群中凋亡细胞的程度。值得注意的是,暴露于GL26CM后,半凝集素-9高的小胶质细胞中凋亡细胞显著增加,但在半凝集素-9阴性/低的小胶质细胞中未检测到这种增加(图4D)。在与GL26-EGFP细胞共培养的原代小胶质细胞中观察到类似的结果(图4E)。为了验证这些结果,作者研究了半乳糖凝集素-9阻断是否可以减少肿瘤暴露的半乳糖凝集素-9高的小胶质细胞的细胞死亡。如图4F所示,半乳糖凝集素-9阻断显著降低了与GL26-EGFP细胞共培养的半乳糖凝集素-9高含量小胶质细胞的死亡细胞百分比,而半乳糖凝集素-9阴性/低含量小胶质细胞没有明显变化(图4F)。总的来说,这些结果表明半乳糖凝集素-9高的小胶质细胞比半乳糖凝集素-9阴性/低的小胶质细胞更容易发生细胞死亡。
5. 半凝集素-9阴性/低的小胶质细胞比半凝集素-9高的小胶质细胞表现出更强的吞噬能力
吞噬作用是中枢神经系统内小胶质细胞的基本功能。因此,作者评估了半乳糖凝集素-9高和半乳糖凝集素-9阴性/低的小胶质细胞对GL26-EGFP细胞的吞噬活性。有趣的是,半乳糖凝集素-9阴性/低的小胶质细胞比半乳糖凝集素-9高的小胶质细胞更有效地吞噬EGFP阳性的肿瘤细胞(图5A)。为了进一步评估半乳糖凝集素-9与小胶质细胞吞噬能力之间的相关性,作者检测了半乳糖凝集素-9表达与代表性抗吞噬分子如PD-1和信号调节蛋白(SIRP)-α的关系。如图5B所示,与PD-1高的小胶质细胞相比,PD-1阴性/低的小胶质细胞对肿瘤细胞的吞噬活性更高。然后作者比较了半乳糖凝集素-9高和半乳糖凝集素-9阴性/低的小胶质细胞中SIRP-α和PD-1的水平。与图5A一致,在与GL26细胞共培养的原代胶质细胞中,半乳糖凝集素-9高的小胶质细胞的SIRP-α和PD-1水平高于半乳糖凝集素-9阴性/低的小胶质细胞(图5C)。为了证实这些结果,作者研究了gl26小鼠肿瘤浸润细胞中SIRP-α和PD-1的水平。半乳糖凝集素-9高cd45midcd11b +的小胶质细胞与半乳糖凝集素-9阴性/低的小胶质细胞相比,显示出更高水平的抗吞噬分子SIRP-α和PD-1(图5D)。在CD45highCD11b+巨噬细胞中观察到类似的结果(图5E)。
接下来,作者研究了半乳糖凝集素-9是否参与与PD-1相关的吞噬作用。为了解决这个问题,作者根据PD-1和半乳糖凝集素-9的表达将CD45midCD11b+小胶质细胞分为四个不同的亚群:PD-1 +Gal-9 -、PD-1 +Gal-9 +、PD-1 - Gal-9 +和PD-1-Gal-9-。然后作者用流式细胞术比较了小胶质细胞亚群中SIRP-α的水平。PD-1 +Gal-9 -和PD-1 +Gal-9 +小胶质细胞在同侧脑半球的比例明显高于对侧脑半球。这些亚群也表现出比其他人群更高的SIRP-α百分比(图5F)。值得注意的是,与PD-1 +Gal-9 -和PD-1 +Gal-9 +小胶质细胞相比,PD-Gal-9 -小胶质细胞中SIRP-α的百分比显著降低。同样,PD-1 +Gal-9 -和PD-1 +Gal-9+小胶质细胞的EGFP+细胞被吞噬的频率较低,而PD-1 - Gal-9 -小胶质细胞的吞噬能力比其他亚群更强(图5G)。
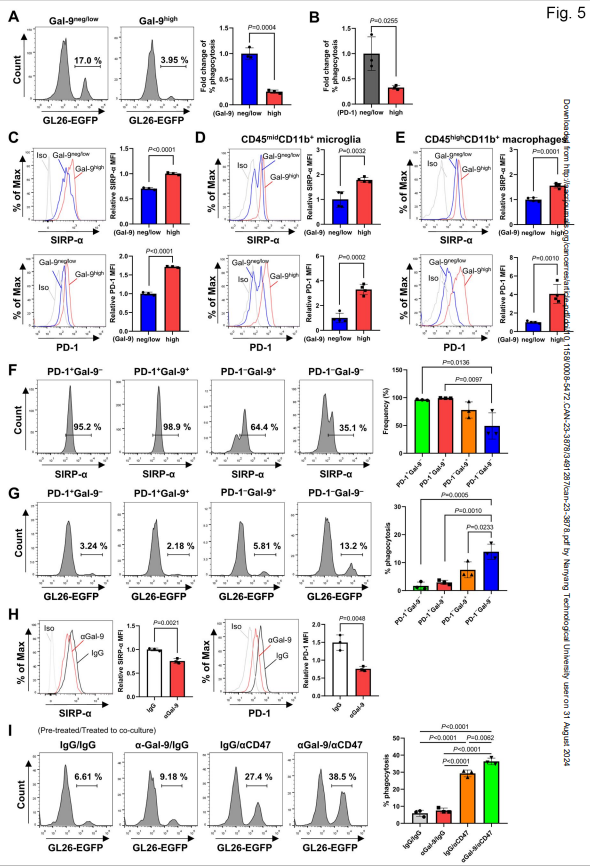
图5 半凝集素-9阴性/低的小胶质细胞比半凝集素-9高的小胶质细胞表现出更强的吞噬能力
然后作者探索半乳糖凝集素-9阻断是否会影响小胶质细胞的吞噬活性。与igg处理的小胶质细胞相比,抗半乳糖凝集素-9抗体处理的原代小胶质细胞显示出更高的头部摄取百分比。此外,在GL26-EGFP细胞培养的原代小胶质细胞中,半凝集素-9阻断可显著降低SIRP-α和PD-1的水平(图5H)。因此,作者研究了半乳糖凝集素-9和CD47的共同阻断是否会影响小胶质细胞肿瘤的吞噬。在原代胶质细胞和GL26-EGFP细胞共培养中,与单独使用任一抗体相比,抗半乳糖凝集素-9和抗cd47抗体共同处理显著增强了吞噬能力。同样,当原代胶质细胞在抗CD47抗体存在下与GL26-EGFP细胞共培养之前,预先暴露于具有抗半乳糖凝集素-9或同型抗体的GL26CM时,双半乳糖凝集素-9和CD47抑制比单次治疗显著增强了小胶质细胞对肿瘤细胞的吞噬作用(图5I)。总之,这些结果支持半乳糖凝集素-9可能有助于脑TME内初级胶质细胞的抗吞噬活性。
6. 阻断半乳糖凝集素-9影响肿瘤大小和脑TME中髓细胞和T细胞的活性
以上结果促使作者进一步研究抗半乳糖凝集素-9抗体对肿瘤生长的影响。GL26细胞移植7天后,腹腔注射200 μg阻断抗体或等量对照IgG;每3天重复一次,持续2周。与IgG对照组相比,半乳糖凝集素-9阻断显著降低了女性和男性肿瘤模型的肿瘤大小,延长了生存期(图6A和B;补充图S16)。肿瘤浸润细胞的FACS分析显示,半乳糖凝集素-9阻断导致CD45midCD11b+小胶质细胞和CD45highCD11b+巨噬细胞中PD-1水平降低(图6C和D)。此外,与igg处理小鼠相比,抗半乳糖凝集素-9处理小鼠的肿瘤内PD-1 +CD8+ T细胞表现出更高的TCF-1表达,TCF-1是CD8+ T细胞效应功能的关键转录因子(图6E)。
先天免疫细胞与T细胞相互作用并触发细胞毒性CD8+ T细胞的激活。因此,作者研究了半乳糖凝集素-9阻断是否会影响胶质细胞和T细胞的相互作用。将原代胶质细胞模拟处理或用GL26CM处理24小时,然后在抗半乳糖凝集素-9抗体或IgG存在下与CD8+ T细胞共培养48小时。流式细胞术显示,抗半乳糖凝集素-9抗体与GL26CM暴露的胶质细胞共培养的PD-1 +CD8+ T细胞中TCF-1水平显著升高(图6F)。ELISA进一步显示,与gl26cm处理的胶质细胞共培养时,抗半乳糖凝集素-9抗体存在时,CD8+ T细胞产生的IFN-α比对照IgG增强(图6G)。与对照IgG相比,半凝集素-9阻断也显著增加了CD107a +脱颗粒CD8+ T细胞的百分比(图6H)。然而,在单独培养的T细胞中,抗半乳糖凝集素-9抗体并不直接影响CD8+ T细胞的活性,包括增殖和细胞因子的产生。这些结果表明,在胶质细胞存在的情况下,半乳糖凝集素-9阻断会影响CD8+ T细胞的活性。
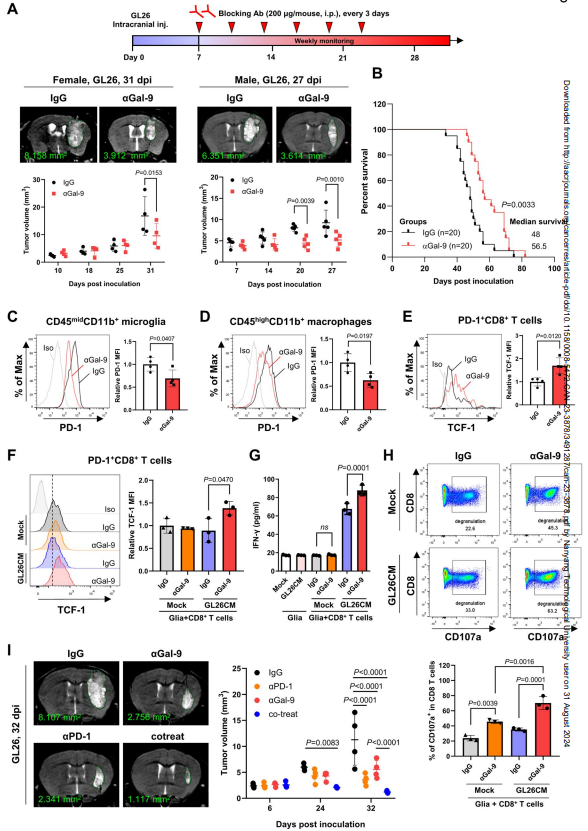
图6 半凝集素-9阻断剂在颅内肿瘤模型中显示抗肿瘤作用
基于以上结果,作者进一步研究抗半乳糖凝集素-9和抗PD -1抗体对肿瘤生长的联合作用。如图6I所示,与IgG对照相比,抗半乳糖凝集素-9或抗PD -1抗体均可显著降低肿瘤体积。值得注意的是,用这两种抗体治疗导致肿瘤大小更明显的缩小。这些结果进一步证实了半凝集素-9阻断在颅内脑肿瘤模型中的作用。
7. Hif-2α对半乳糖凝集素-9的表达和脑TME内肿瘤的生长至关重要
作者接下来的问题是半乳糖凝集素-9的表达是如何在脑TME的胶质细胞中被调节的。鉴于小鼠半乳糖凝集素-9基因的启动子区域包含2个假定的缺氧反应元件(HRE;5 ' -(A/G)CGT(G/C)(G/C)-3 ')在其启动子区域与缺氧诱导因子(Hif)(33)结合(Supplementary Fig. S20A),作者研究了半凝集素-9在脑TME中的表达是否受到Hif的调节。使用吡莫硝唑(hypoxyprobe-1),一种缺氧标志物,作者发现半乳糖凝集素-9在同侧半球肿瘤边缘的吡莫硝唑染色区域表达更为普遍(图7A),与上述结果一致,免疫细胞化学和流式细胞术显示,在1%氧气缺氧条件下,小胶质细胞中半凝集素-9的表达明显高于20%氧气正常缺氧条件下的小胶质细胞(图7B和C)。
为了进一步研究小胶质细胞半凝集素-9的表达是否受到缺氧的主要调节因子hif的调节,作者使用了髓细胞特异性Hif-1α-或Hif-2α-缺陷小鼠。作者收集gl26接种的LysM-Cre、LysM-Cre+/+/Hif-1α f/f和LysM-Cre+/+/Hif-2α f/f小鼠脑组织,比较半凝集素-9水平。在LysM-Cre和LysM-Cre+/+/Hif-1α f/f小鼠的组织中,半凝集素-9在同侧区域的表达明显高于对侧区域(图7D)。值得注意的是,Western blotting和免疫荧光染色显示,与LysM-Cre小鼠相比,LysM-Cre+/+/Hif-2α f/f小鼠同侧组织中半乳糖凝集素-9水平明显降低(图7D)。流式细胞术进一步显示,在LysM-Cre小鼠中,肿瘤浸润性CD45midCD11b+小胶质细胞上的半凝集素-9表达显著升高,但在LysM-Cre+/+/Hif-2α f/f小鼠中,这种升高明显降低(图7E)。此外,MRI和免疫组织化学分析显示,雄性和雌性LysM-Cre+/+/Hif-2α f/f小鼠的肿瘤体积明显小于LysM-Cre小鼠(图7F)
为了进一步证明Hif-2α依赖性半凝集素-9在小胶质细胞中的表达,作者使用原代胶质细胞进行了体外实验。与LysM-Cre对照小鼠相比,LysM-Cre+/+/Hif-2α f/f小鼠gl26cm暴露的胶质细胞中半乳糖凝集素-9水平明显降低(图7G)。此外,GL26CM对LysM-Cre小鼠小胶质细胞中半乳糖凝集素-9和PD-L1转录物的上调在LysM-Cre+/+/Hif-2α f/f小鼠中未见(图7H)。与小胶质细胞类似,敲除星形胶质细胞中的Hif-2α可显著抑制半乳糖凝集素-9的表达。此外,即使在正常条件下,GL26CM和rmGal-9也能提高Hif-2α转录水平。这些数据强烈表明,Hif-2α在调节脑TME内胶质细胞的半乳糖凝集素-9中起着重要作用。

图7 Hif-2α调控脑TME中半凝集素-9的表达
结论:
基于这些临床相关结果和作者的发现,作者提出半乳糖凝集素-9作为小胶质细胞的关键免疫检查点分子,影响脑TME的免疫和炎症反应。这个研究结果表明,将半乳糖凝集素-9靶向于髓系细胞可能是一种有希望的策略,可以改善基于icb的免疫治疗,克服脑肿瘤的治疗无反应性和耐药性。
实验方法:
颅内脑肿瘤模型,脑浸润细胞的分离,原代小胶质细胞的培养,条件培养基的制备,免疫印迹和RT-PCR分析,免疫荧光成像,流式细胞术,程序性细胞死亡实验,胶质细胞与CD8+ T细胞共培养实验
参考文献:
Chanju Lee, Dahee Yu, Hyung-Seok Kim, Ki Sun Kim, Chi Young Chang, Hee Jung Yoon, Su Bin Won, Dae Yeon Kim, Eun Ah Goh, Yong Sun Lee, Jong-Bae Park, Sang Soo Kim, Eun Jung Park; Galectin-9 Mediates the Functions of Microglia in the Hypoxic Brain Tumor Microenvironment. Cancer Res 15 November 2024; 84 (22): 3788–3802. https://doi.org/10.1158/0008-5472.CAN-23-3878