






m7G修饰的mt-tRF3b-LeuTAA通过软骨细胞中SIRT3的SUMO化来调节线粒体自噬和代谢重编程
骨关节炎(OA)是一种退行性疾病,软骨细胞退化是其发展的关键。越来越多的证据表明,m7G RNA修饰与OA病理过程相关,但其在软骨细胞退化中的具体作用尚不明确。在该研究中,作者发现,在骨关节炎软骨细胞中,METTL1 和 m7G的水平显著升高。此外,METTL1介导的m7G修饰会上调线粒体转运 RNA 衍生片mt-tRF3b-LeuTAA的表达,从而加剧软骨细胞的退变。从机制上讲,mt-tRF3b-LeuTAA 会降低 SUMO 特异性蛋白酶 1(SENP1)的蛋白表达,并上调去乙酰化酶 3(SIRT3)的 SUMO 化水平,进而抑制 PTEN 诱导激酶 1(PINK1)/ 帕金蛋白(Parkin)介导的线粒体自噬。关节腔内注射聚酰胺-胺-聚乙二醇表面修饰的含最小自肽和软骨细胞亲和肽的mt-tRF3b-LeuTAA抑制剂(PMC)可在体内减轻内侧半月板失稳(DMM)小鼠软骨的退变。作者的研究表明,METTL1/m7G/mt-tRF3b-LeuTAA 轴通过SIRT3的SUMO化抑制线粒体自噬并促进线粒体功能障碍,从而加速软骨降解,并且提示靶向METTL1及其下游信号轴可能是骨关节炎治疗的一个有前景的治疗靶点。该研究于2024年10月发表在《Biomaterials》上,IF:12.8。
技术路线:
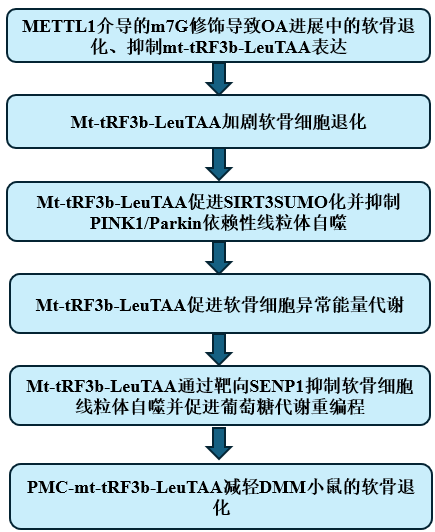
主要研究结果:
1.METTL1介导的m7G修饰导致OA进展中的软骨退化
为了研究m7G甲基转移酶修饰在软骨退化中的作用,作者通过RNA-seq技术检查了m7G调节剂的表达。作者发现METTL1表达在OA软骨细胞中显著升高(图1A和B)。此外,基质金属蛋白酶13(MMP13)表达和m7G修饰水平在OA软骨细胞中显著增加,而胶原蛋白II型alpha1(COL2A1)表达显著降低(图1C和D,n=30)。Kellgren-Lawrence等级用于评估软骨样本的退化等级(KLG;0-4)。结果显示METTL1表达与OA进展中的m7G修饰水平呈显著相关性(图1E)。
为了进一步验证METTL1介导的m7G修饰是否会加速OA的病理进展,将METTL1过表达质粒或METTL1-sh质粒转染到原代人软骨细胞中。作者观察到METTL1过表达显著上调了MMP13表达并下调了COL2A1表达。同时,m7G的总水平显著增加(图1F-H)。此外,功能获得和丧失实验表明,过表达和沉默METTL1均显著影响软骨细胞增殖、凋亡以及合成代谢和分解代谢标志物的表达(图1I和J)。总之,这些发现表明METTL1介导的m7G修饰有助于OA中的软骨细胞退化。
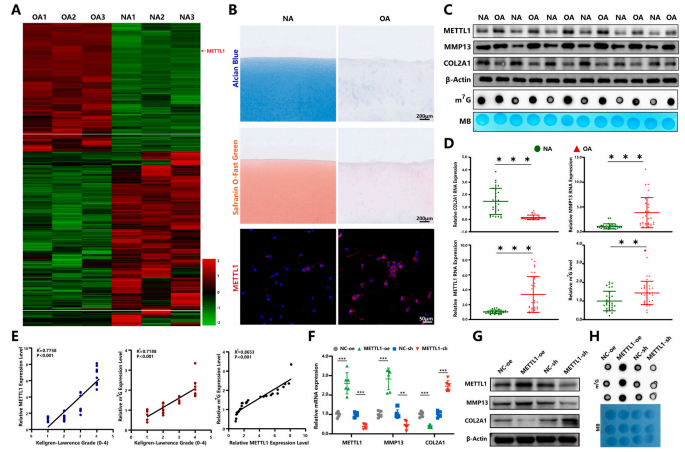
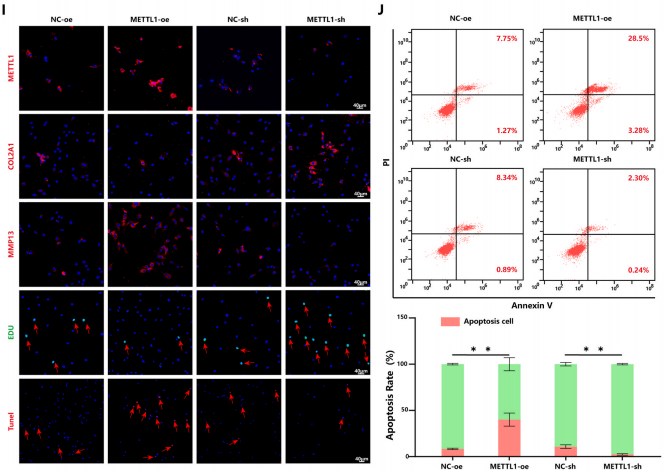
图 1. OA软骨细胞中的METTL1表达和m7G修饰水平促进软骨细胞退化
2.METTL1介导的m7G修饰抑制mt-tRF3b-LeuTAA表达
m7G是一种常见的tRNA衍生片段修饰,在原核生物、真核生物和一些古细菌中高度保守。为了进一步探索METTL1介导的m7G修饰的具体机制,进行了arraystar人类m7G小RNA表观转录组微阵列。作者注意到软骨细胞中许多tRF具有m7G修饰(图2A-C)。在临床样本中筛选并验证了六种具有高m7G修饰的tRF。其中,与正常软骨细胞相比,mt-tRF3b-LeuTAA表达在OA软骨细胞中更稳定且更高(图2D)。mt-tRF3bLeuTAA源自mt-tRNALeuTAA,其源tRNA序列和结构是从MINT基础数据库中获得的(图2D)。
Pearson相关性分析显示mt-tRF3b-LeuTAA表达与m7G修饰、METTL1水平和KLG之间存在很强的相关性(图2E)。此外,m7G修饰RNA免疫沉淀表明mt-tRF3b-LeuTAA在m7G位点高度富集(图2F)。METTL1敲低上调了mttRF3b-LeuTAA的表达,这与METTL1过表达形成对比(图2G)。免疫荧光(IF)和荧光原位杂交(FISH)显示METTL1和mt-tRF3b-LeuTAA在软骨细胞中共定位(图2H和I)。以上结果表明mt-tRF3bLeuTAA上存在m7G修饰,而METTL1是该修饰的原因。
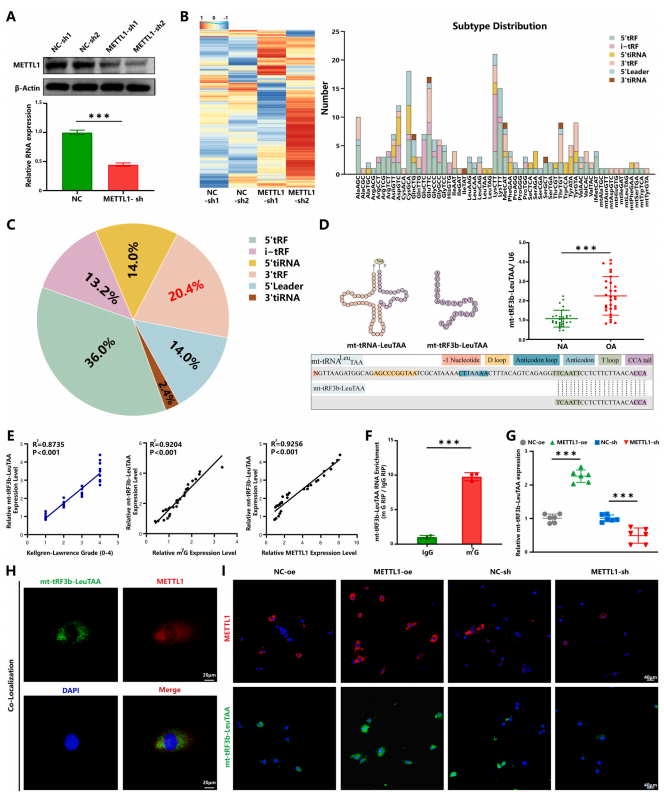
图2. METTL1介导的m7 G-甲基化tRNA衍生片段的鉴定
3.Mt-tRF3b-LeuTAA加剧软骨细胞退化
为了进一步研究mt-tRF3b-LeuTAA在OA中的作用,将mt-tRF3bLeuTAA模拟物、mt-tRF3b-LeuTAA抑制剂和对照转染到软骨细胞中。IF结果显示,mt-tRF3bLeuTAA模拟物增加了软骨细胞中MMP13的表达并降低了COL2A1的表达。然而,mt-tRF3b-LeuTAA抑制剂产生了相反的效果。此外,mt-tRF3b-LeuTAA模拟物D.LongetalBiomaterials314(2025)122903通过EdU测定抑制了软骨细胞增殖,并通过流式细胞术和TUNEL测定增加了软骨细胞凋亡。Westernblot分析显示,mttRF3b-LeuTAA模拟物上调了软骨细胞外基质中分解代谢标志物(MMP13、RUNX2和ADAMTS4)的水平,并降低了软骨相关合成代谢标志物(Aggrecan、SOX9和COL2A1)的水平。这些结果表明mt-tRF3b-LeuTAA导致OA中的软骨退化。
tRF可以通过靶向mRNA来抑制基因转录。作者利用生物信息学数据库(miRanda、TargetScan和tRFTars)预测并筛选了mt-tRF3b-LeuTAA靶向的6个mRNA,包括TNF受体相关因子2(TARF2)、甲状腺激素受体相互作用蛋白4(TRIP4)、C-X-C基序趋化因子配体8(CXCL8)、细胞周期蛋白D2(CCND2)、ras相关蛋白Rab-2A(RAB2A)和sentrin特异性蛋白酶1(SENP1)。在软骨细胞中过表达mt-tRF3bLeuTAA后,只有SENP1表达显著降低,这表明SENP1是mt-tRF3b-LeuTAA的有效下游靶点。这些发现表明mt-tRF3b-LeuTAA可能通过靶向SENP1促进软骨细胞退化。
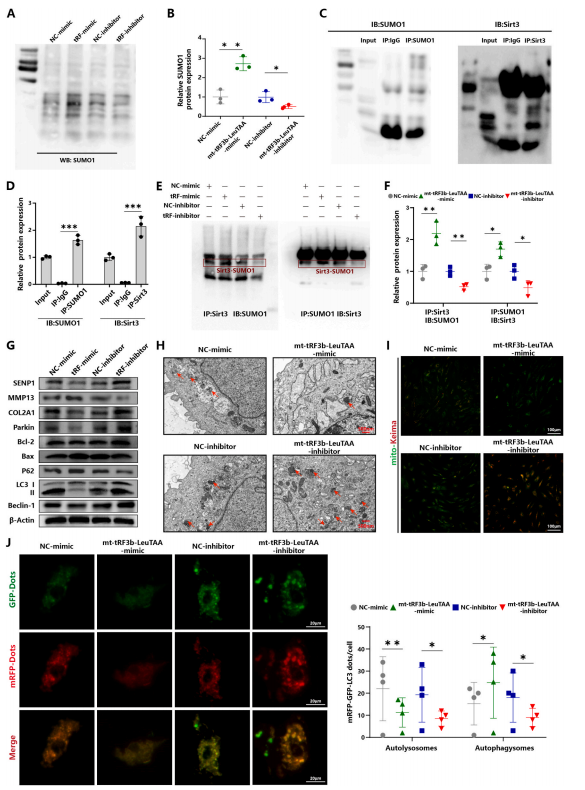
图 3. mt-tRF3b-LeuTAA 通过 PINK1/Parkin 通路促进 SIRT3 SUMO 化,从而抑制软骨细胞线粒体自噬
4.Mt-tRF3b-LeuTAA促进SIRT3SUMO化并抑制PINK1/Parkin依赖性线粒体自噬
作者之前的研究表明,SIRT3通过PINK1/Parkin通路激活线粒体自噬。其他研究表明,SIRT3含有1型SUMO化(SUMO1)位点,SUMO1修饰可通过降低底物去乙酰化活性来抑制SIRT3功能。为了验证mt-tRF3b-LeuTAA是否通过PINK1/Parkin通路影响SIRT3SUMO化和线粒体自噬,在原代人软骨细胞中敲低或过表达mt-tRF3b-LeuTAA。作者发现mt-tRF3b-LeuTAA模拟物导致总SUMO1修饰更高,而mt-tRF3b-LeuTAA抑制剂具有相反的效果(图3A和B)。免疫共沉淀实验证实mt-tRF3b-LeuTAA模拟物增强了SIRT3SUMO1修饰(图3C-F)。为进一步探讨mt-tRF3b-LeuTAA调控软骨细胞线粒体自噬的机制,作者进行了Westernblot、mRFP-GFP-LC3双标腺病毒及mito-Keima实验。结果显示mt-tRF3b-LeuTAA模拟物抑制了软骨细胞线粒体自噬相关蛋白Parkin、LC3II/I、Beclin-1及COL2A1,增加了P62及MMP13。此外,mt-tRF3b-LeuTAA模拟物增加了凋亡相关蛋白BAX,抑制了增殖相关蛋白Bcl-2(图3G)。mRFP-GFP-LC3、透射电子显微镜(TEM)和线粒体-凯马分析显示,mt-tRF3b-LeuTAA模拟物下调了软骨细胞中的线粒体自噬通量(图3H-J)。这些发现表明,mt-tRF3b-LeuTAA促进SIRT3SUMO化并抑制OA中的PINK1/Parkin依赖性线粒体自噬。
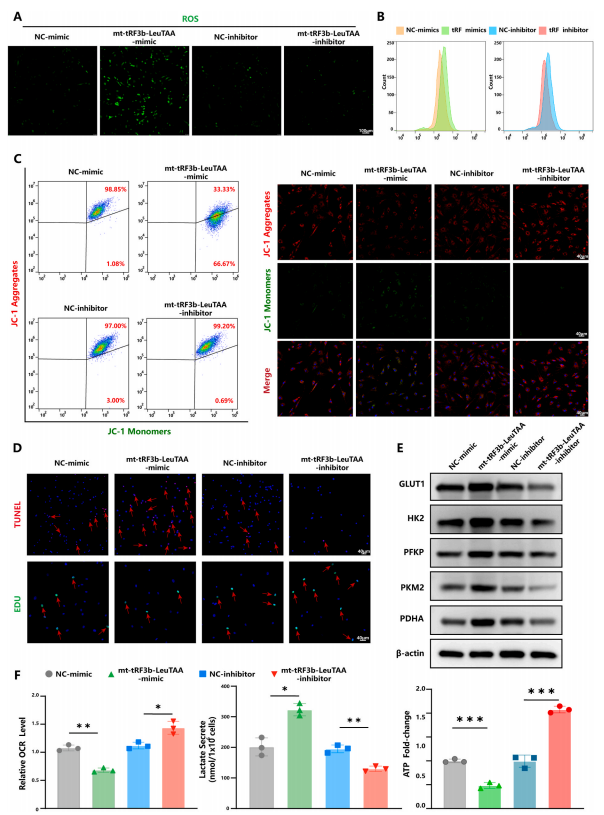
图 4. mt-tRF3b-LeuTAA 通过线粒体自噬调节软骨细胞葡萄糖代谢重编程
5.Mt-tRF3b-LeuTAA促进软骨细胞异常能量代谢
线粒体自噬也与能量代谢有关,包括氧化磷酸化 (OXPHO) 和糖酵解。为了确定mt-tRF3b-LeuTAA是否调节软骨细胞中的OXPHO和糖酵解,对线粒体的代谢功能进行了评估。作者发现mttRF3b-LeuTAA模拟了活性氧(ROS)生成增加(图4A和B),以及线粒体膜电位降低、软骨细胞增殖和细胞凋亡增加(图4C和D)。Westernblot分析显示,mttRF3b-LeuTAA模拟物可导致糖代谢相关酶水平升高,包括葡萄糖转运蛋白1型(GLUT1)、己糖激酶2(HK2)、磷酸果糖激酶(PFKP)、丙酮酸激酶肌同工酶M2(PKM2)和丙酮酸脱氢酶E1(PDHA)(图4E)。此外,mt-tRF3b-LeuTAA模拟物可降低耗氧率(OCR)水平,增加乳酸生成,减少ATP生成(图4F)。然而,在软骨细胞中敲低mt-tRF3b-LeuTAA则会引起相反的效果。这些结果证实,mt-tRF3b-LeuTAA下调线粒体自噬并促进糖酵解,破坏正常的能量代谢,导致OA中持续的ROS和乳酸沉积。
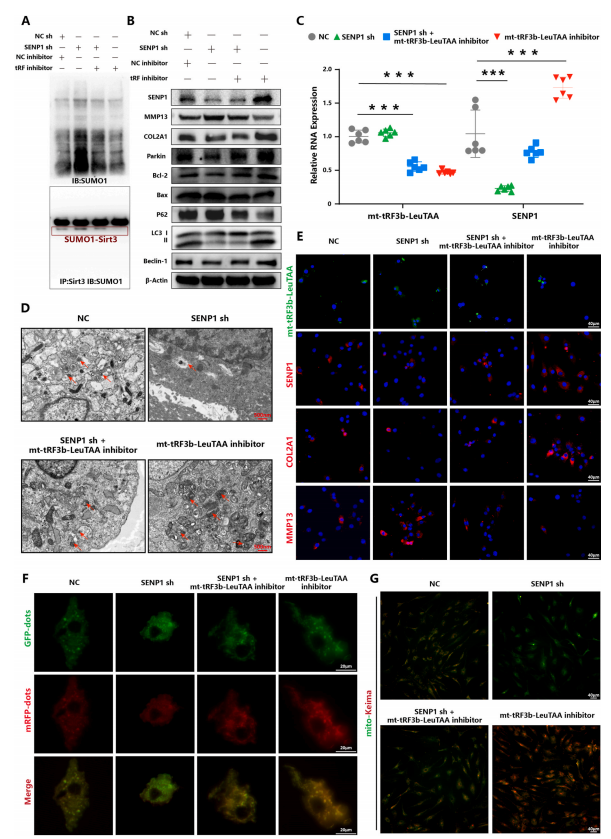
图 5. mt-tRF3b-LeuTAA 通过靶向 SENP1 抑制软骨细胞线粒体自噬
6.Mt-tRF3b-LeuTAA通过靶向SENP1抑制软骨细胞线粒体自噬并促进葡萄糖代谢重编程
SENP1作为SUMO1修饰的主要调控酶,对SIRT3SUMO1修饰具有较高的水解效率。为了确认mt-tRF3b-LeuTAA是否通过靶向SENP1增强SIRT3SUMO1修饰并抑制PINK1/Parkin介导的线粒体自噬,作者进行了一系列实验。首先,将mt-tRF3b-LeuTAA模拟物和mt-tRF3b-LeuTAA抑制剂转染到原代人软骨细胞中。作者发现mt-tRF3b-LeuTAA模拟物显著抑制了SENP1的表达,这与mt-tRF3b-LeuTAA抑制剂形成对比。
然后,将mt-tRF3b-LeuTAA和SENP1分别或同时转染到软骨细胞中。结果表明,SENP1敲低增强了总SUMO1修饰和SIRT3SUMO1修饰(图5A)。此外,SENP1敲低通过检测mRFP-GFP-LC3、mito-Keima和线粒体自噬相关蛋白表达水平抑制了PINK1/Parkin介导的线粒体自噬(图5B-G)。同时,作者还发现SENP1敲低降低了线粒体膜电位,增加了ROS生成(图6A-D),并促进了糖酵解(图6E和F)。重要的是,通过转染mt-tRF3b-LeuTAA抑制剂可以挽救SENP1敲低的影响。此外,SENP1过表达检测到了相反的效果,而通过用mt-tRF3b-LeuTAA模拟物转染可以挽救这些效果,进一步支持SENP1作为mt-tRF3b-LeuTAA的功能介质。最后,作者通过Targetscan和miRbase数据库预测了mt-tRF3b-LeuTAA和SENP1的潜在结合位点。作者发现mt-tRF3bLeuTAA与SENP1mRNA的3′-非翻译区(UTR)结合。此外,双荧光素酶报告基因测定证实mttRF3b-LeuTAA可以直接结合SENP1mRNA3′-UTR。
总体而言,这些结果表明mt-tRF3b-LeuTAA靶向SENP1来调节SIRT3SUMO1修饰,从而抑制PINK1/Parkin介导的线粒体自噬并促进软骨细胞中的葡萄糖代谢重编程。
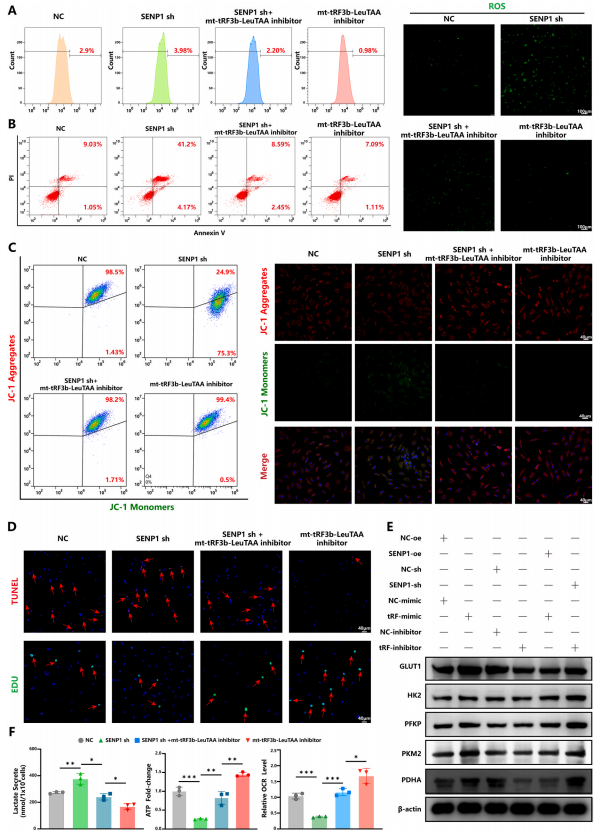
图6. mt-tRF3b-LeuTAA 通过靶向SENP1 促进软骨细胞代谢重编程
7构建针对软骨细胞的高亲和力纳米载体
基于上述mt-tRF3b-LeuTAA调控软骨细胞稳态的机制,作者进一步探索mt-tRF3b-LeuTAA在OA中的治疗潜力。首先,构建软骨细胞亲和性及吞噬细胞逃逸性纳米粒子,以聚酰胺胺(PAMAM)为载体,将最小“自身”肽(MSP)和软骨细胞亲和肽(CAP)附着于纳米载体上(图7A),将该纳米载体体系命名为PMC(Polyamidoamine-polyethyleneglycolsurface-modifiedwithMinimalself-peptidesandChondrocyte-affinitypeptides)。游离miRNA形成的亮带在PMC处完全消失,tRF的重量比为0.5–14,表明tRF分子已被PMC复合和阻滞。琼脂糖凝胶电泳表明,在N/P比为8时,所有tRF分子均未复合(图7B)。细胞毒性试验表明,在40-50%聚乙二醇(PEG)的纳米胶囊对原代人软骨细胞的毒性较低,而RT-qPCR测定发现PEG比例为40%的纳米胶囊可提供更佳的递送效果(图7C和D)。
然后,将mt-tRF3b-LeuTAA装载到PMC中。平均粒径为45.8nm,正电荷为10.7mV(图7E-G)。为了评估PMC的降解和持续释放,将OA滑液与PMC-mt-tRF3b-LeuTAA和裸露的mt-tRF3b-LeuTAA一起孵育。RNA扩增和琼脂糖凝胶电泳显示与PMC培养24h后可检测到条带(图7H)。
接下来,将人软骨外植体与PMC-mt-tRF3bLeuTAA(FITC)或Nakedmt-tRF3b-LeuTAA(FITC)一起培养24h,进行体外渗透实验。结果显示PMC-mt-tRF3b-LeuTAA组荧光增强,而Nakedmt-tRF3b-LeuTAA组呈现相反趋势(图7I)。同样,将巨噬细胞和软骨细胞与PMC-mttRF3b-LeuTAA或Nakedmt-tRF3b-LeuTAA一起培养24h。在PMC-mt-tRF3bLeuTAA组中,软骨细胞比巨噬细胞表现出更强的荧光。(图7J)。然后,作者通过将PMC-mt-tRF3b-LeuTAA添加到六种不同类型的细胞中来研究PMC的靶向软骨细胞能力。结果表明,与其他类型的细胞相比,软骨细胞特异性地吸收了PMC-mt-tRF3b-LeuTAA。最重要的是,PMC-mt-tRF3b-LeuTAA是一种很有前途的纳米载体,可用于进一步评估OA的体内治疗效果。
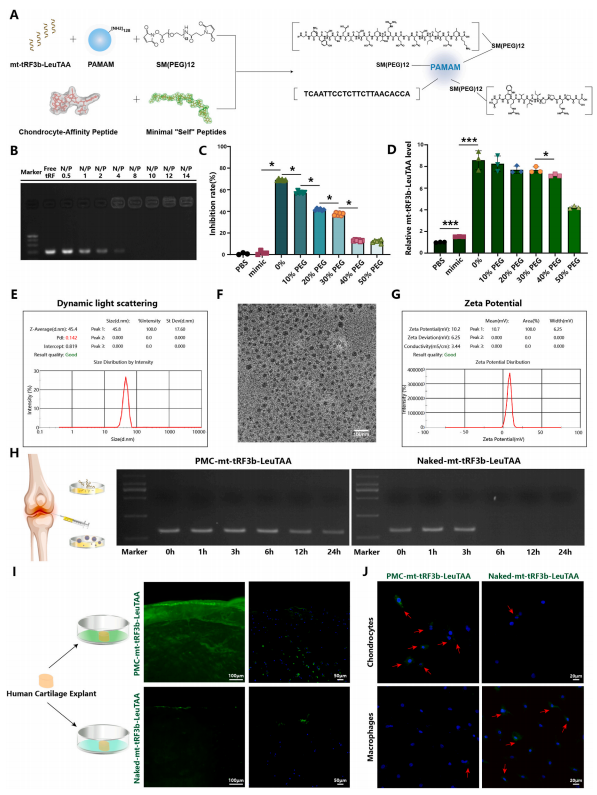
图 7. 软骨细胞亲和吞噬细胞逃逸纳米粒子促进 mt-tRF3b-LeuTAA 递送到软骨细胞中
8.PMC-mt-tRF3b-LeuTAA减轻DMM小鼠的软骨退化
为了验证PMC-mt-tRF3b-LeuTAA在体内治疗OA的效果,合成了Cy5.5标记的mt-tRF3b-LeuTAA,并制备了相应的纳米递送材料和裸RNA对照。对雄性C57BL/6小鼠进行关节内注射。三周后,器官采集和成像显示PMC-mt-tRF3b-LeuTAA和裸RNA对照组之间没有显著差异,表明PMC-mt-tRF3b-LeuTAA在体内的毒性很小。体内成像显示裸mt-tRF3b-LeuTAA的荧光随时间下降的速度比PMC-mt-tRF3b-LeuTAA更快。
然后对雄性C57BL/6小鼠进行内侧半月板失稳术(DMM),建立OA模型。所有小鼠随机分为Sham组、DMM组、DMM+Nakedmt-tRF3b-LeuTAA组、DMM+PMC-mttRF3b-LeuTAA组和DMM+PMC-mt-tRF3b-LeuTAA抑制剂组。DMM+Control组小鼠注射Nakedmt-tRF3bLeuTAA,DMM+PMC组小鼠注射PMCmt-tRF3b-LeuTAA或DMM+PMC-mt-tRF3b-LeuTAA抑制剂。术后10周,软骨关节间隙、层表面、基质染色、软骨细胞排列、细胞萎缩、反应性聚集、软骨下骨血管增生等均有明显差异(图8A)。免疫组织化学和免疫荧光显示DMM+PMC-mt-tRF3b-LeuTAA组的MMP13、COL10A1、RUNX2表达较高,COL2A1和Aggrecan表达较低,而DMM+PMC-mt-tRF3b-LeuTAA抑制剂组则呈现相反趋势(图8B)。小鼠膝关节的微型计算机断层扫描评估了骨赘和硬化半月板。与单纯DMM组相比,DMM+PMC-mt-tRF3b-LeuTAA组的骨赘体积和等级较高,而DMM+PMCmt-tRF3b-LeuTAA抑制剂组检测到的骨赘较低(图8C)。这些发现意味着PMC-mt-tRF3b-LeuTAA抑制剂可减轻DMM小鼠的软骨退化。综上所述,PMC可以有效地将tRF递送到软骨细胞中,为OA提供一种有前景的基于纳米技术的精准靶向策略。
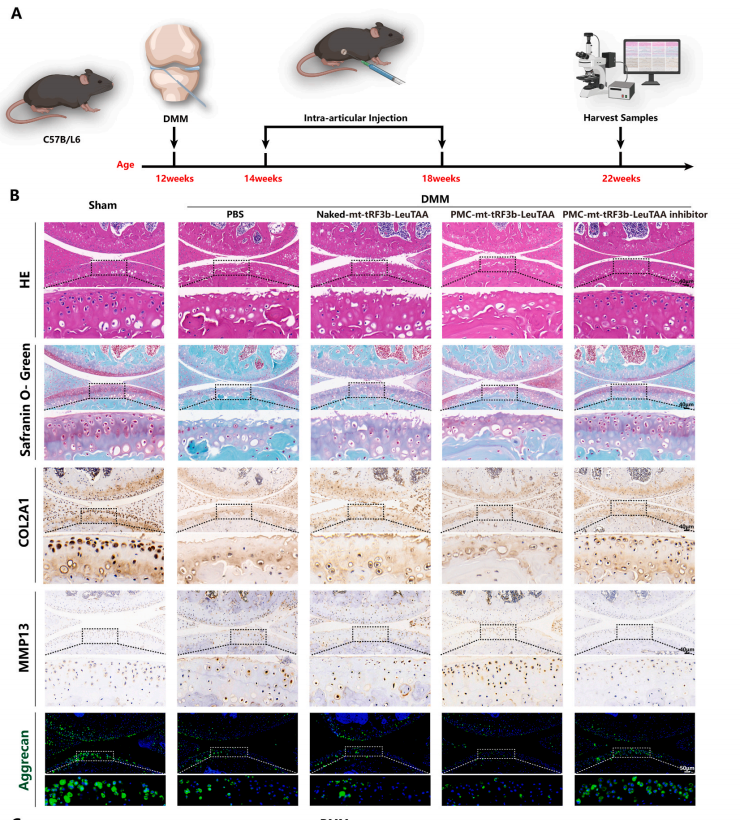
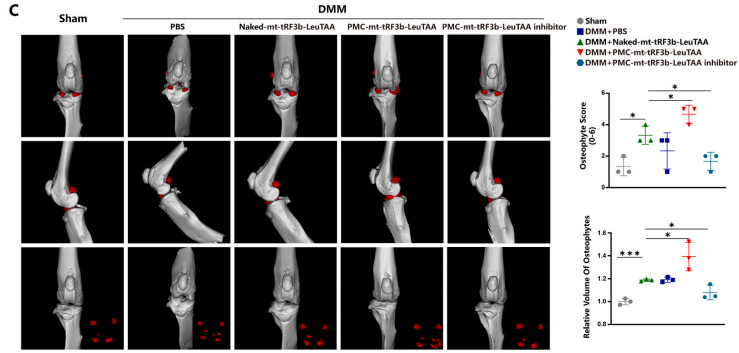
图 8. PMC-mt-tRF3b-LeuTAA 抑制剂减轻 DMM 小鼠的软骨退化
结论:
综上所述,METTL1 介导的m7 G修饰上调了mt-tRF3b-LeuTAA的表达,而mt-tRF3bLeuTAA则通过靶向SENP1来促进SIRT3 SUMO1修饰,从而抑制 SIRT3的功能。该过程抑制了软骨细胞线粒体自噬,促进了糖酵解和代谢重编程,并加速了软骨细胞的退化。在转化应用中,mt-tRF3bLeuTAA代表了研究线粒体自噬和OA进展的新靶点。此外,作者合成了一种新型纳米载体PMC,可显著增强tRF进入软骨细胞,并为OA中的tRF特异性核酸药物提供了一种新的治疗策略。
参考文献:
Long D, Deng Z, Zhao X, et al. m7G-modified mt-tRF3b-LeuTAA regulates mitophagy and metabolic reprogramming via SUMOylation of SIRT3 in chondrocytes. Biomaterials. Published online October 23, 2024. doi:10.1016/j.biomaterials.2024.122903
实验方法:
组织病理学染色、qPCR、WB、Dot blot、RT-qPCR、免疫荧光染色、流式细胞术、荧光原位杂交、TUNEL、 CCK-8、micro-CT、骨关节炎小鼠模型的构建