肝细胞癌(HCC)是全世界最常见的癌症之一。5-甲基胞嘧啶(m5C)RNA甲基转移酶NSUN2参与细胞增殖和转移,并在多种癌症中上调。然而,NSUN2介导的m5C修饰在HCC中的生物学功能和调控机制尚未得到很好的研究。我们的结果显示,在接受肝切除的HCC患者中,NSUN2表达上调并与预后不良相关。NSUN2过表达显著促进了HCC的生长和转移,而NSUN2敲减则产生了相反的效果。m5C RNA免疫沉淀测序(m5C-RIP-Seq)揭示,m5C超甲基化与mRNA过表达相关,并且NSUN2介导的m5C超甲基化促进了HCC患者的代谢。从机制上讲,我们的数据揭示PKM2是NSUN2介导的m5C修饰的下游靶标。具体来说,NSUN2可以通过增加PKM2 mRNA的3′-UTR中C773位点的m5C水平来稳定PKM2 mRNA。此外,拯救实验表明,NSUN2通过上调PKM2促进了HCC的糖酵解和进展。总之,本研究揭示了NSUN2介导的m5C修饰通过稳定PKM2 mRNA促进了肝细胞癌的糖酵解和进展,并为HCC患者提供了潜在的预后因素和治疗靶点。本文于2025年2月发表于“Cell Death Dis”(IF=8.1)上。
技术路线:

结果:
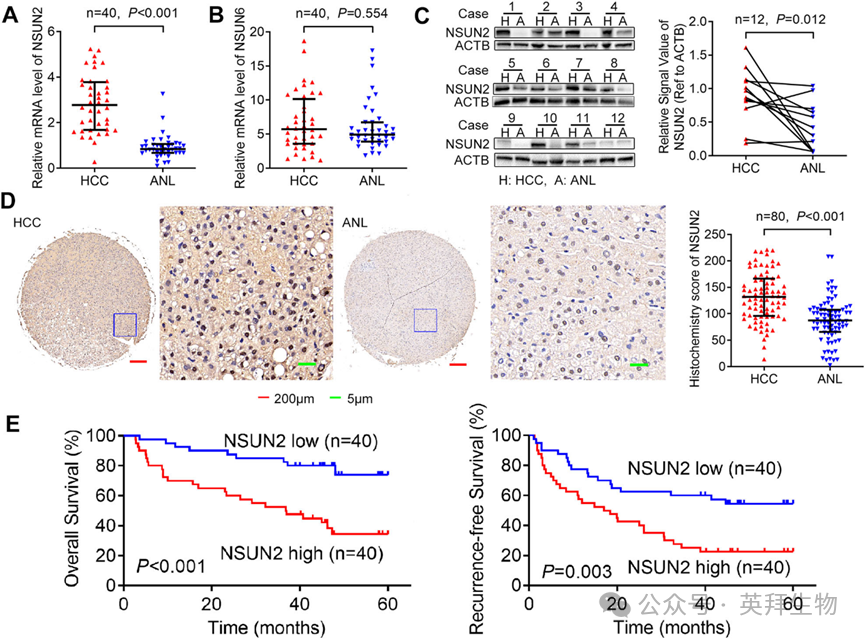
1)在肝切除术后HCC患者中,NSUN2表达上调并与预后不良相关
为了确定mRNA m5C修饰在HCC中的作用,我们通过RT-qPCR(队列1)评估了40对HCC和ANL(非肝细胞癌肝脏组织)组织中两种主要的mRNA m5C甲基转移酶NSUN2和NSUN6的表达水平。结果显示,NSUN2的mRNA水平在HCC中增加(图1A),而NSUN6的mRNA水平没有显著差异(图1B)。此外,对从队列1中随机选取的12对HCC和ANL组织进行western blotting分析,发现NSUN2的蛋白水平在HCC中也上调(图1C)。为了确定高NSUN2表达在HCC中的临床重要性,我们通过免疫组化(IHC)(图1D)在另一个具有预后数据的80名HCC患者队列(队列2)中检测了NSUN2的表达。此外,NSUN2蛋白在HCC中过表达。根据HCC组织中NSUN2的IHC评分中位数,我们将80名HCC患者分为高NSUN2表达组和低NSUN2表达组。Kaplan-Meier生存曲线显示,在接受肝切除的HCC患者中,NSUN2的高表达与较低的总体生存率(OS)和较低的复发无病生存率(RFS)相关(图1E)。
2)NSUN2在体外和体内促进HCC的生长和转移
然后,我们探讨了NSUN2是否影响HCC的进展。CCK-8和Transwell实验表明,强制表达NSUN2促进了HCC的生长和转移(图2A, B),而NSUN2的耗竭抑制了HCC的生长和转移(图2C, D)。为了探索NSUN2在体内HCC生长中的作用,将NSUN2过表达的SNU387细胞及其相应的阴性对照细胞,以及NSUN2沉默(NSUN2-sh2)的Hep3B细胞及其相应的阴性对照细胞皮下注射到裸鼠中。每周评估肿瘤体积,直到4周后处死裸鼠。NSUN2过表达显著促进了SNU387细胞的生长,而NSUN2敲减抑制了Hep3B细胞的生长(图2E, F)。此外,为了确定NSUN2在体内HCC转移中的作用,我们通过将NSUN2过表达或NSUN2沉默的HCC细胞注射到裸鼠的尾静脉中,建立了肺转移模型。使用体内成像系统监测HCC细胞在肺中的转移过程,持续5周。结果显示,NSUN2的上调增加了HCC细胞的肺转移,而NSUN2的下调减少了肺转移(图2G, H)。总的来说,NSUN2能够在体外和体内诱导HCC的生长和转移。

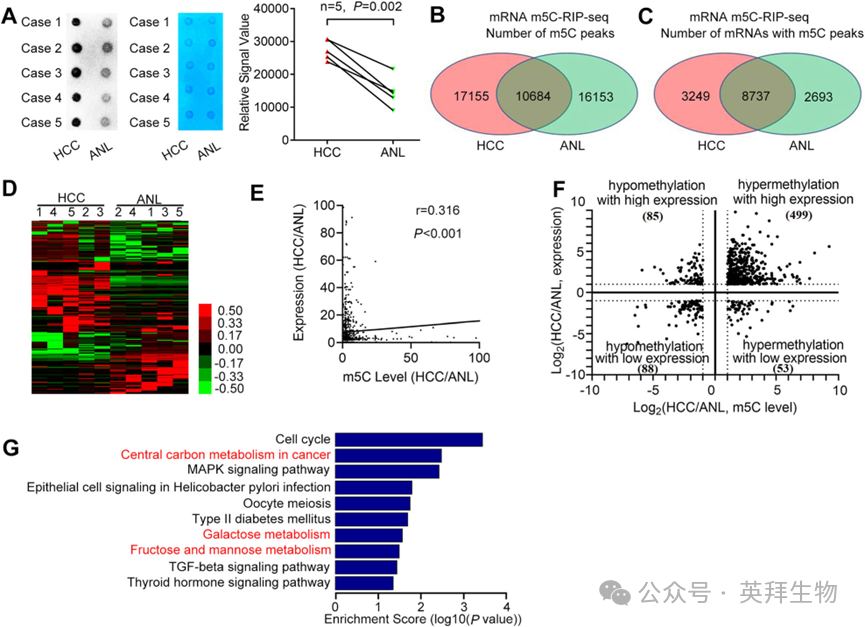
3)NSUN2介导的m5C高甲基化促进HCC的代谢
接下来,我们探讨了NSUN2在HCC中的致癌作用是否依赖于m5C。首先,为了检测人类HCC中的全局mRNA m5C水平,我们在五对HCC和ANL组织(队列3)中进行了m5C点杂交实验(图3A)。HCC中的全局mRNA m5C水平高于ANL组织。此外,我们对这五对HCC和ANL组织的mRNAs进行了m5C-RIP-seq,以阐明HCC的转录组m5C谱。其中,有10684个m5C峰在8737个mRNAs中同时在HCC和ANL组织中检测到(图3B, C)。比较了HCC和ANL中mRNAs的m5C水平,确定了在HCC中上调的3156个甲基化峰在2696个mRNAs中,而在HCC中检测到下调的1998个甲基化峰在1756个mRNAs中(图3D)。总的来说,这些发现表明m5C在HCC中的mRNAs经常出现过甲基化。mRNA m5C-RIP-seq和mRNA-Seq数据的联合分析显示,HCC中mRNAs的表达与它们的m5C水平略微正相关(图3E)。此外,在3156个上调的m5C峰中,对应的499个(15.81%)mRNAs在HCC中表达上调,而只有53个(1.68%)在HCC中表达下调(图3F)。在1998个下调的m5C峰中,对应的85个(4.25%)mRNAs在HCC中表达上调,88个(4.40%)在HCC中表达下调。这些结果表明,m5C超甲基化与HCC中mRNA的过表达正相关。KEGG通路分析显示,既表达上调又m5C水平上调的mRNAs在十个通路中富集(图3G)。有趣的是,癌症中的中心碳代谢(HK2, MAPK3, MYC, PFKP, PKM2, SLC16A3)、半乳糖代谢(B4GALT2, HK2, PFKP)和果糖及甘露糖代谢(HK2, PFKFB4, PFKP)相关的通路都与代谢有关。总的来说,m5C超甲基化与HCC中的mRNA过表达和代谢相关。
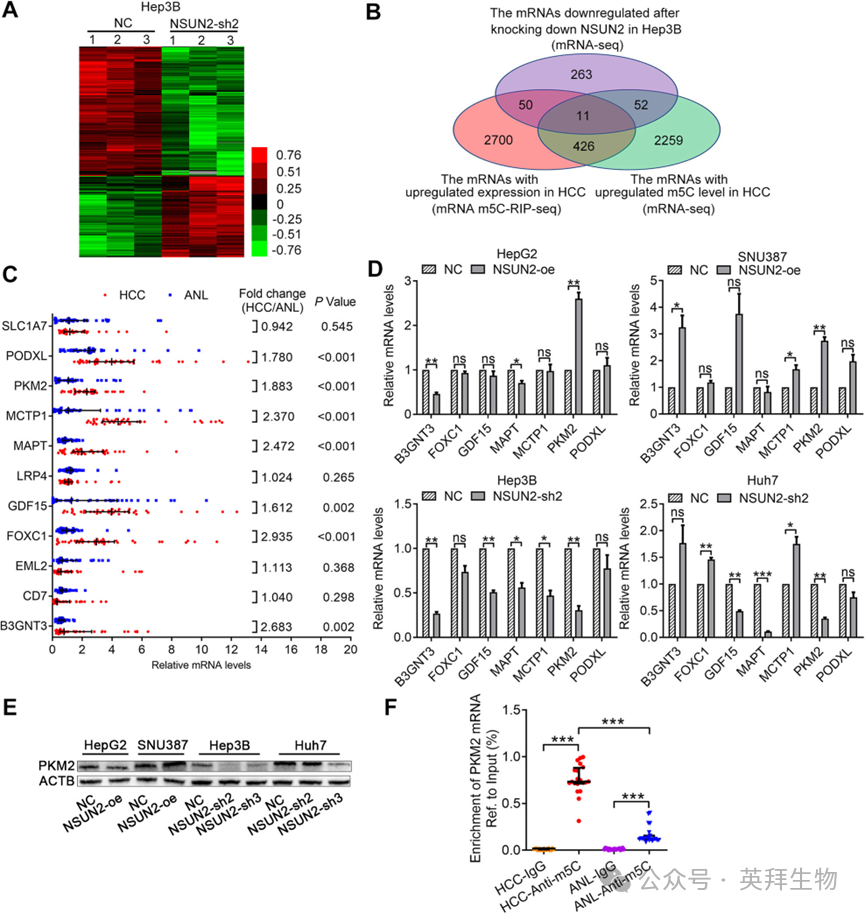
4)PKM2是NSUN2介导的m5C修饰促进代谢的主要靶点
为了识别人类HCC中NSUN2的靶mRNA,我们对Hep3B-NSUN2-sh2细胞及其相应的阴性对照细胞进行了mRNA测序。在敲减NSUN2后,有236个基因表达上调,376个基因表达下调(图4A)。然后,我们将HCC中表达上调的mRNAs、HCC中m5C水平上调的mRNAs以及Hep3B细胞中NSUN2敲减后表达下调的mRNAs进行重叠。维恩图显示,有十一个mRNAs(B3GNT3, CD7, EML2, FOXC1, GDF15, LRP4, MAPT, MCTP1, PKM2, PODXL和SLC1A7)符合这些条件(图4B, C)。我们接着在40对HCC和ANL组织中检测了这十一个mRNAs的表达,并发现其中七个在HCC中表达上调(图4C)。此外,我们在HCC细胞系中检测了这七个mRNAs的表达,发现只有PKM2在HepG2和SNU387细胞中过表达NSUN2后上调,而在Hep3B和Huh7细胞中沉默NSUN2后下调(图4D, E)。根据m5C-RIP-seq的结果,PKM2 mRNA上调的m5C峰位于其3′-UTR(chr15:72491753-72491855, hg19)。针对这个峰的m5C-RIP-qPCR验证了这些结果(图4F)。因此,我们假设PKM2 mRNA是NSUN2的主要靶标。
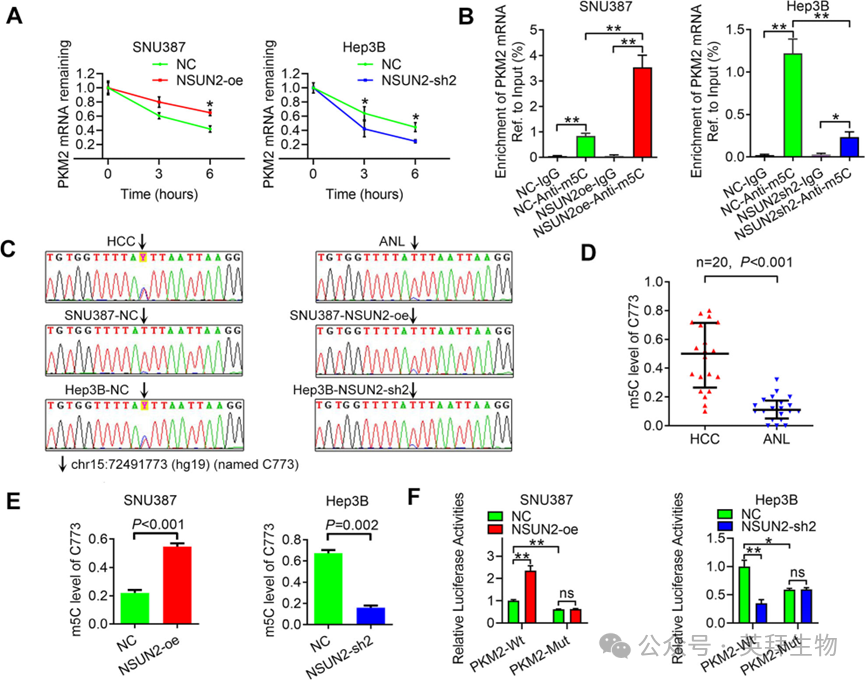
5)NSUN2通过增加其m5C水平来稳定PKM2 mRNA
接下来,我们研究了NSUN2是否可以通过增加其m5C水平来稳定PKM2 mRNA,从而诱导PKM2的表达。我们发现,过表达NSUN2减缓了PKM2 mRNA的降解,而沉默NSUN2则产生了相反的效果(图5A)。这些发现确认了NSUN2可以稳定PKM2 mRNA。此外,m5C-RIP-qPCR显示,在SNU387细胞中过表达NSUN2后,PKM2 mRNA的m5C水平增加,而在Hep3B细胞中敲减NSUN2后,m5C水平降低(图5B)。为了检测PKM2的确切m5C位点,我们使用针对PKM2 m5C峰的引物和从亚硫酸盐转化RNA逆转录的cDNA模板进行了亚硫酸盐-PCR。代表性的Sanger测序色谱图显示,chr15:72491773(hg19)(命名为C773)的信号与胞嘧啶(‘C’,蓝色)和胸腺嘧啶(‘T’,红色)混合,表明HCC组织和细胞系中m5C被甲基化(图5C)。使用先前描述的方法,我们计算了每个样本中的m5C水平。例如,从SNU387-NC的PCR产物中亚克隆并插入T-easy载体,随机选择了50个克隆并进行测序。其中,9个克隆被检测为‘C’,而41个克隆被检测为‘T’。因此,SNU387-NC中C773的m5C水平计算为18%(9/50)。每个样本检测三次。使用这种方法,也检测了其他样本中C773的m5C水平(图5D–F)。HCC组织中的m5C水平高于ANL组织(图5D)。此外,在SNU387细胞中过表达NSUN2后,m5C水平增加,而在Hep3B细胞中敲减NSUN2后,m5C水平降低(图5E)。为了验证C773在调节PKM2表达中的作用,我们使用带有野生型PKM2 mRNA的3′-UTR的质粒(PKM2-WT)或带有突变C773的质粒(PKM2-Mut)进行了荧光素酶报告基因实验。正如预期的那样,强制表达NSUN2增加了PKM2-WT的荧光素酶活性,但并未增加PKM2-Mut的活性(图5F),而沉默NSUN2则抑制了这种活性,表明NSUN2对PKM2的调节作用依赖于m5C位点C773。总之,NSUN2可以通过增加PKM2 mRNA的3′-UTR中m5C位点C773的m5C水平来稳定PKM2 mRNA。
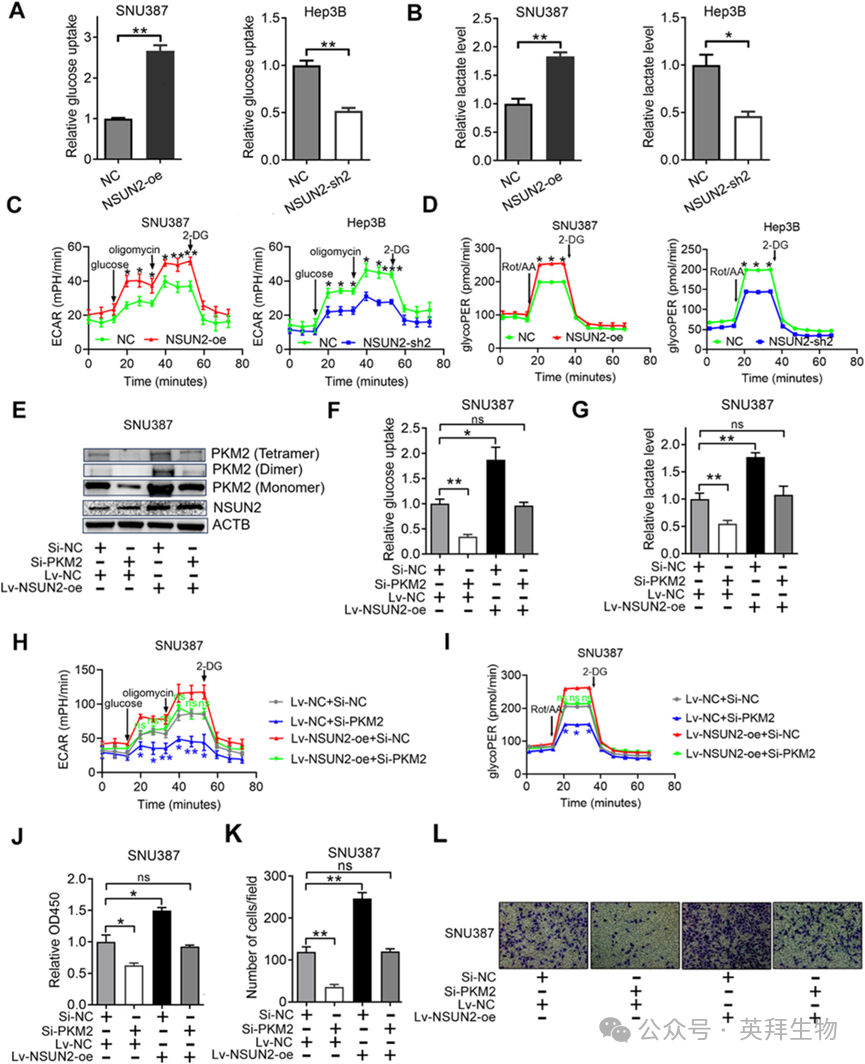
6)NSUN2通过上调PKM2促进HCC糖酵解和进展
Warburg效应的特点是葡萄糖摄取增加和乳酸产生增加。为了测试NSUN2是否可以通过增强PKM2介导的Warburg效应来促进HCC的进展,我们在过表达和敲减NSUN2后检测了葡萄糖摄取和乳酸产生。我们发现,过表达NSUN2显著增加了HCC细胞中的葡萄糖摄取和乳酸产生,而沉默NSUN2则减少了HCC细胞中的葡萄糖摄取和乳酸产生(图6A, B)。重要的是,NSUN2过表达增加了ECAR,而NSUN2沉默则减少了HCC细胞的ECAR(图6C)。为了检查过表达和沉默NSUN2后细胞的糖酵解能力,还进行了GlycoPER的测定。结果显示,过表达NSUN2导致HCC细胞中的GlycoPER显著增加。此外,沉默NSUN2导致HCC细胞中的GlycoPER减少(图6D)。我们接下来研究了NSUN2对HCC的影响是否是通过PKM2介导的。首先,我们发现针对PKM2的siRNA可以下调PKM2四聚体、二聚体和单体(四聚体、二聚体和单体的表达比例没有变化)的表达,而同时过表达NSUN2可以减缓这种效果(图6E)。其次,我们发现沉默PKM2抑制了葡萄糖摄取和乳酸产生,这种效果被过表达NSUN2所阻断(图6F, G)。此外,沉默PKM2降低了ECAR和GlycoPER,而同时过表达NSUN2则减缓了这种效果(图6H, I)。另外,敲减PKM2抑制了HCC细胞的生长和侵袭,这种效果被过表达NSUN2所阻断(图6J–L)。总之,NSUN2通过上调PKM2促进了HCC的糖酵解和进展。
结论:
我们的研究揭示了NSUN2在HCC进展中的关键作用,并证实这种作用依赖于它的m5C修饰。体外和体内实验进一步证实了NSUN2促进HCC生长和转移的作用。机制上,NSUN2介导的m5C修饰通过稳定PKM2 mRNA促进糖酵解和HCC的进展。这些发现提示NSUN2介导的m5C修饰在HCC中发挥重要作用,为HCC患者提供了潜在的预后因素和治疗靶点。
实验方法:
KEGG通路分析、mRNA表达与m5C修饰相关性分析、m5C-RIP-seq、mRNA-Seq、基因表达分析、基因编辑与过表达、m5C点印迹、m5C-RIP-qPCR、荧光素酶报告基因实验、亚硫酸盐-PCR、细胞培养与处理、细胞增殖与迁移实验、细胞代谢检测、小鼠肿瘤模型、病理分析
参考文献:
Qi Q, Zhong R, Huang Y, Tang Y, Zhang XW, Liu C, Gao CF, Zhou L, Yu J, Wu LY. The RNA M5C methyltransferase NSUN2 promotes progression of hepatocellular carcinoma by enhancing PKM2-mediated glycolysis. Cell Death Dis. 2025 Feb 9;16(1):82. doi: 10.1038/s41419-025-07414-5.






