






ELK3-CYFIP2轴介导的肌动蛋白重塑调节三阴性乳腺癌的转移和自然杀伤细胞反应
三阴性乳腺癌(TNBC)是一种侵袭性、高转移性疾病,预后较差。E26转化特异性转录因子(ELK3)在TNBC中高度表达,是上皮-间质转化和免疫反应的调节因子。由于TNBC细胞的转移迁移和免疫逃避是成功转移的关键因素,因此揭示其潜在机制并开发有效的免疫治疗策略迫在眉睫。本研究通过RNA测序分析和分子实验,确定并验证与TNBC细胞迁移和附着相关的ELK3下游靶基因。致癌基因ELK3直接抑制细胞质FMR1相互作用蛋白2(CYFIP2)的表达,而CYFIP2是肌动蛋白聚集的抑制因子。进一步的分子和药理学分析证实,ELK3-CYFIP2 轴在 TNBC 细胞系中具有双重作用:(1)通过调节肌动蛋白的积累控制丝状体介导的迁移和粘附;(2)通过调节接触部位的肌动蛋白积累调节对 NK 细胞的敏感性。Kaplan-Meier分析表明,ELK3-CYFIP2轴与TNBC患者的存活率有关,ELK3抑制CYFIP2的转录。因此,ELK3-CYFIP2轴在调节肌动蛋白方面起着关键作用,强调了它在控制TNBC癌细胞迁移和NK细胞反应方面的重要性。该研究于2025年2月发表在《Journal of Experimental & Clinical Cancer Research》,IF:11.4。
技术路线:

主要研究结果:
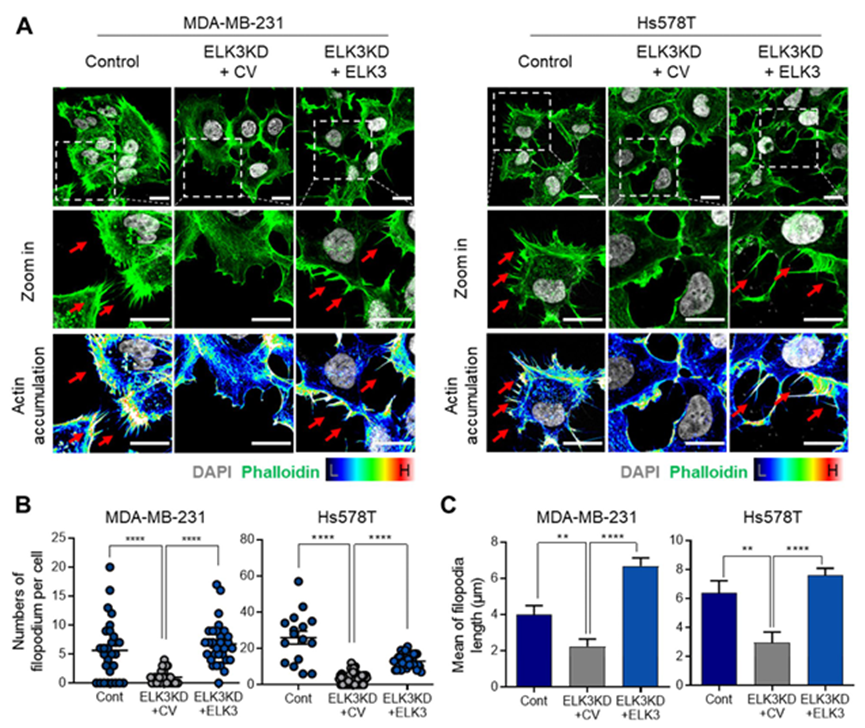
1. ELK3 的表达与 TNBC 细胞的丝状突起有关
丝状体是从细胞表面延伸出来的细长的指状突起,在癌细胞的运动、迁移和转移中起着至关重要的作用。据此,高度转移的 MDA-MB-231 细胞在体外表现出许多类似丝状体的结构。鉴于 ELK3-knockdown(ELK3KD)MDA-MB-231 细胞失去其转移特性,作者质疑 ELK3KD 细胞是否具有较少的丝状体。为证实这一点,暂时恢复ELK3KD MDA-MB-231或Hs578T细胞中的ELK3表达。不出所料,用类黄嘌呤染色观察肌动蛋白丝时发现,ELK3KD 细胞比对照细胞表现出更少的细胞表面突起;然而,恢复 ELK3 表达后,细胞表面表现出更多的突起(图 1A 左)。在另一种 TNBC 细胞系 Hs578T 细胞中也观察到类似的模式(图 1A 右)。图 1A 中的定量分析表明,ELK3 表达与每个 MDA-MB-231 和 Hs578T 细胞丝状突起的数量和平均长度之间存在显著相关性(图 1B-C)。
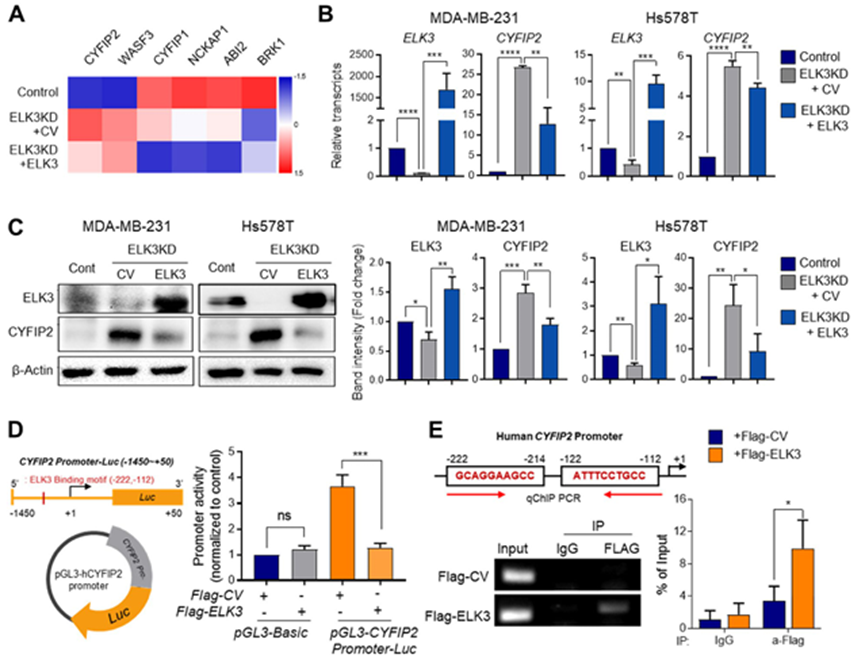
2. ELK3 在 TNBC 中发挥着 CYFIP2 转录抑制因子的功能
丝状结构由 WAVE 复合物(即 WASF,又称 WAVE)、NCKAP1、ABI 和 BRICK1(或其同源物)以及连接膜和细胞骨架的 CYFIP 组成。由于ELK3在TNBCs中起转录抑制作用,通过分析MDA-MB-231细胞、ELK3KD细胞和ELK3-rescued ELK3KD细胞的RNA测序数据,评估编码各WAVE成分的基因中哪些基因与ELK3的表达呈负相关。
虽然在ELK3KD细胞中CYFIP2和WASF3的表达受到抑制,但挽救ELK3在ELK3KD细胞中的表达并没有显著抑制WASF3/WAVE3的表达。这表明,ELK3可能并不像调控CYFIP2那样直接调控WASF3/WAVE3。基于这一点,重点关注 CYFIP2,因为在 RNA 测序数据中,它与 ELK3 的表达呈明显的负相关(图 2A)。CYFIP2 是肌动蛋白聚合的负调控因子,可抑制丝状体的形成。与图 2A 中显示的结果一致,定量分析显示,在 ELK3KD 的 MDA-MB-231 和 Hs578T 细胞中,CYFIP2 mRNA 和蛋白上调,而当 ELK3 恢复后,它们的表达量减少(图 2B,C)。在CYFIP2的启动子上靠近转录起始位点的地方发现一个ELK3结合基团(图1)。为证明CYFIP2是ELK3在TNBCs中的直接下游靶点,采用荧光素酶报告试验,使用编码荧光素酶的构建体,该构建体携带CYFIP2启动子区域的bp -1450至+50。如图 2D 所示,ELK3 显著抑制 CYFIP2 启动子的活性。通过 ChIP 检测证实 ELK3 与 CYFIP2 启动子的直接结合,结果显示 Flag-ELK3 与相应的基团结合(图 2E)。这些发现表明,ELK3在TNBCs中发挥着CYFIP2转录抑制因子的功能。
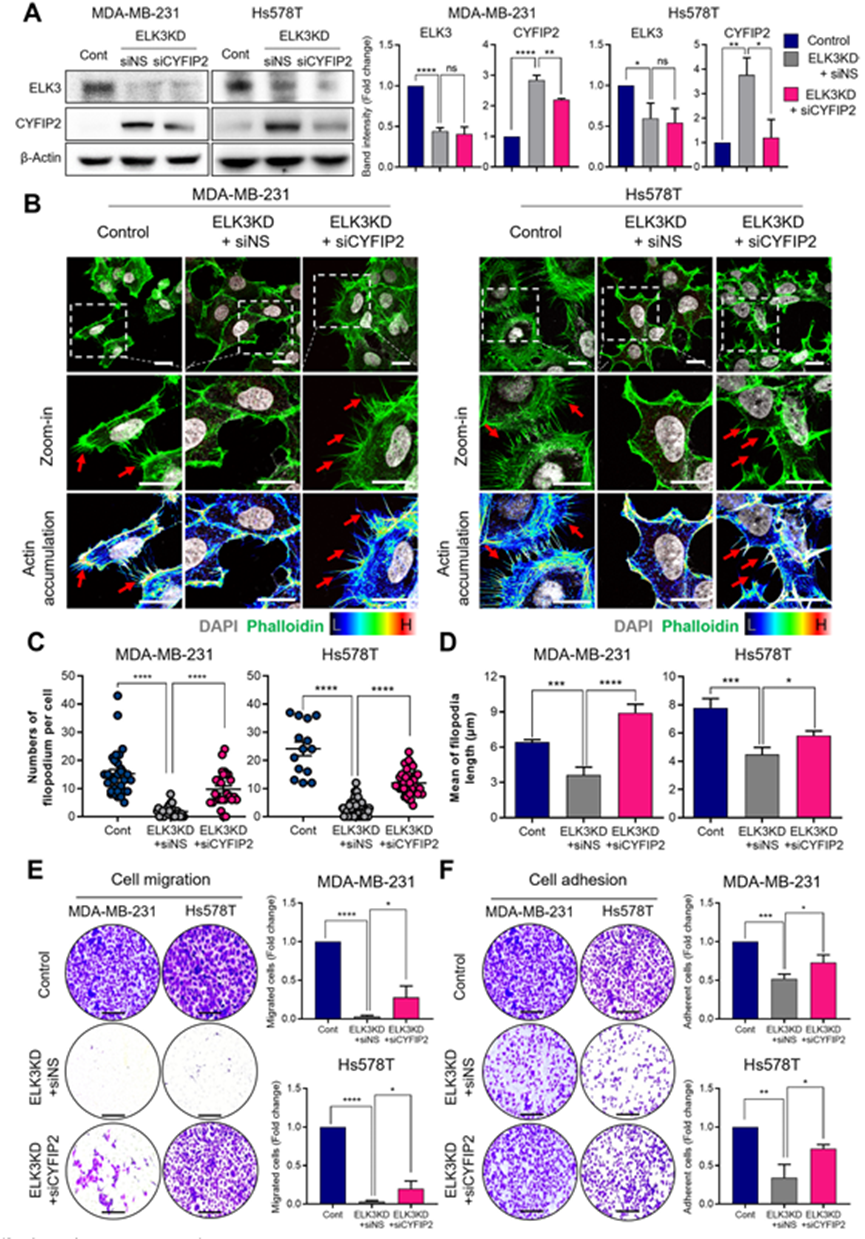
3. ELK3-CYFIP2轴通过调节丝状突起调控TNBC的转移特性
为进一步探索ELK3-CYFIP2轴在TNBCs中的生物学功能,作者了评估抑制CYFIP2对ELK3KD TNBCs转移特性的影响。通过将 siRNA 转染 ELK3KD MDA-MB-231 和 Hs578T 细胞,成功抑制CYFIP2(图 3A)。值得注意的是,CYFIP2的抑制导致ELK3KD MDA-MB-231和Hs578T细胞的细胞表面突起重新出现,每个细胞的丝状突起平均数量和平均长度的增加就是证明(图3B-D)。
此外,抑制 ELK3KD 的 MDA-MB-231 和 Hs578T 细胞中的 CYFIP2 可增强其迁移能力,而 ELK3 的抑制会削弱这种能力(图 3E)。当 CYFIP2 被抑制时,在 ELK3KD 细胞中观察到的低细胞粘附性增加(图 3F)。这些结果表明,ELK3-CYFIP2轴调节TNBCs的关键转移特性,包括丝状体的形成、迁移和粘附。
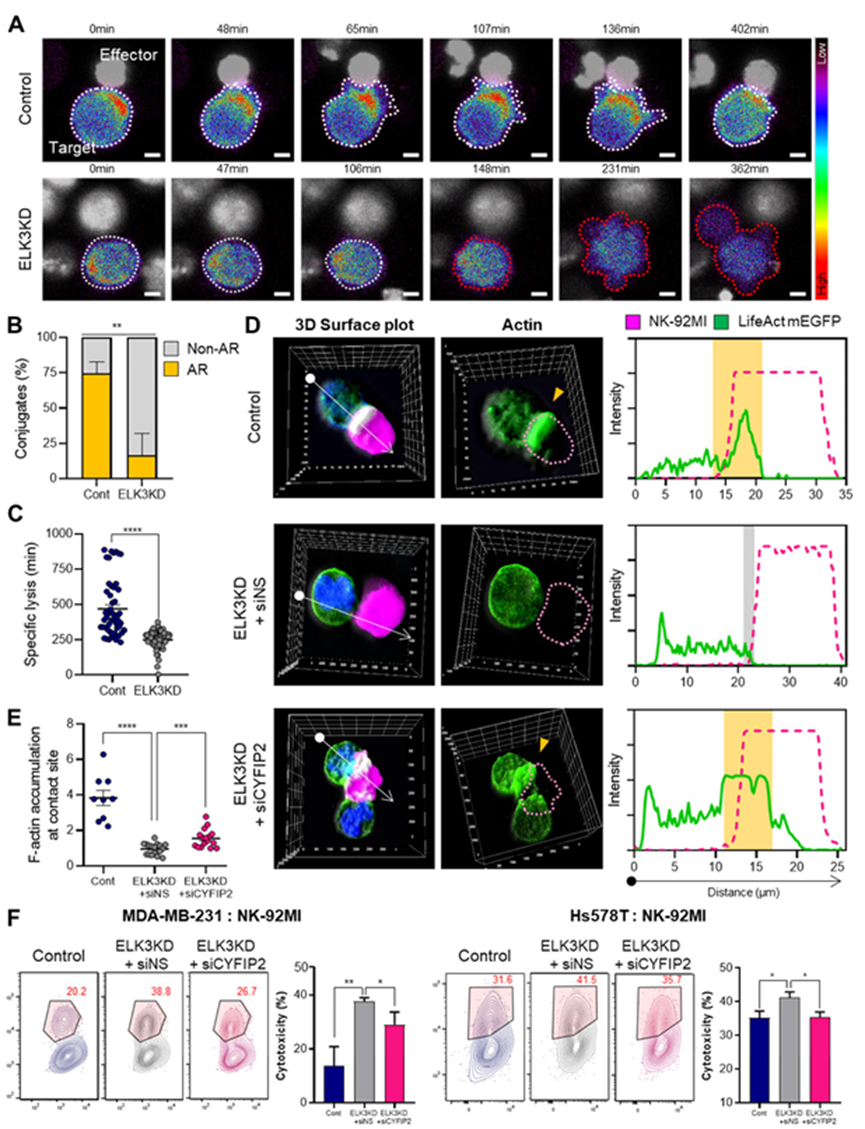
4. ELK3-CYFIP2轴通过调节肌动蛋白积聚影响TNBC对NK细胞的敏感性
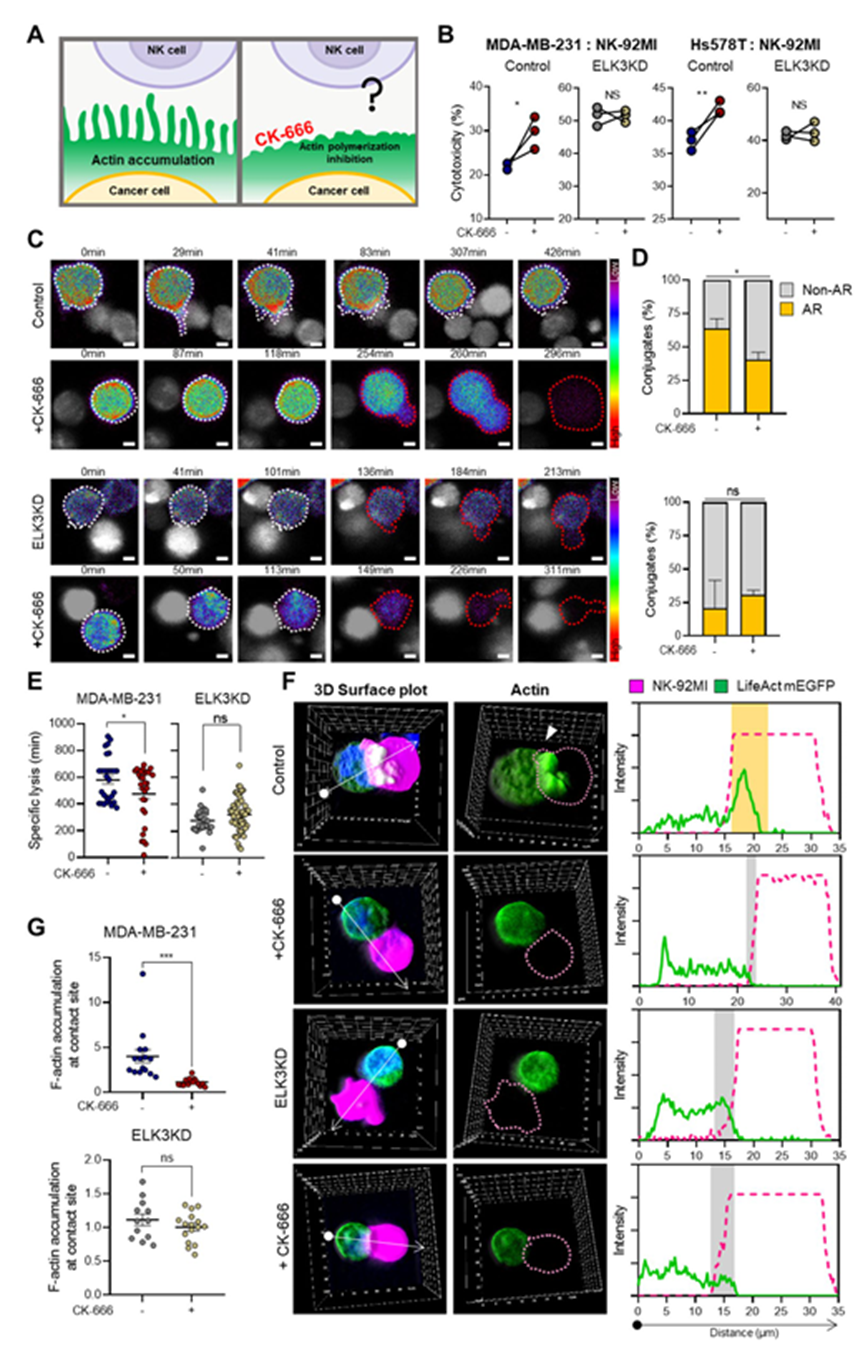
乳腺癌细胞中的肌动蛋白积聚可诱导细胞抵抗 NK 细胞介导的细胞毒性。在与 NK-92MI 细胞共培养的 MDA-MB-231 对照细胞和 ELK3KD 细胞中观察融合GFP 的肌动蛋白,发现对照细胞与 ELK3KD 细胞接触部位的肌动蛋白堆积突出(图 4A)。对与 NK 细胞接触部位显示 GFP 融合肌动蛋白聚集的癌细胞进行定量,进一步证实 ELK3KD 细胞与 NK 细胞接触后肌动蛋白聚集减少的观察结果(图 4B)。此外,ELK3KD 细胞被 NK 细胞杀死的时间明显缩短(图 4C),表明肌动蛋白反应影响细胞毒性。为证实 TNBCs 中的 ELK3-CYFIP2 轴是否参与针对 NK 细胞的免疫反应过程中的肌动蛋白积累,将 siCYFIP2 转染到 ELK3KD TNBCs 中。抑制 CYFIP2 增加了 ELK3KD 细胞与 NK 细胞接触时的肌动蛋白堆积(图 4D-E)。同样,图 4F 显示转染 siCYFIP2 的 ELK3KD-TNBC 中 NK 细胞毒性明显降低。这些结果突显了 ELK3-CYFIP2 轴通过肌动蛋白聚集在这些癌细胞对 NK 细胞的免疫敏感性中的作用。
5. 药物抑制肌动蛋白重塑对 MDA-MB-231 细胞对 NK 细胞免疫反应的影响
上述数据表明,ELK3KD细胞与NK细胞接触部位的肌动蛋白组装因CYFIP2的表达上调而受损,从而增加它们对NK细胞的敏感性。根据CYFIP2诱导WAVE复合物失稳的报道,作者推测,在ELK3KD TNBCs中,作用于WAVE下游的Arp2/3受损。为进一步证实这一点,接下来研究在对NK细胞免疫敏感的情况下,ELK3KD细胞是否对肌动蛋白重塑化学抑制剂有反应。
CK-666 通过抑制 Arp2/3 破坏肌动蛋白细胞骨架动力学和肌动蛋白重塑(图 5A),对 ELK3KD MDA-MB-231 和 Hs578T 细胞对 NK 细胞的免疫反应没有影响,但显著增强对照细胞的敏感性(图 5B)。延时显微镜观察发现,CK-666抑制NK细胞接触部位对照细胞的丝状突起,而ELK3KD MDA-MB-231细胞对这种化学物质没有反应(图5C)。定量分析证实,CK-666 对 ELK3KD 细胞肌动蛋白对 NK 细胞接触的反应没有影响(图 5D)。与CK-666对MDA-MB-231和ELK3KD细胞肌动蛋白积累的影响一致,CK-666缩短NK细胞杀死MDA-MB-231对照细胞的时间,但不影响杀死ELK3KD细胞的时间(图5E)。接下来评估CK-666 对癌细胞与 NK-92MI 细胞接触部位肌动蛋白相对堆积的影响。CK-666处理后,MDA-MB-231细胞的GFP融合肌动蛋白积累减少,而ELK3KD细胞则无明显变化(图5F-G)。这些结果表明,在TNBCs的ELK3KD中,Arp2/3已经失去活性,导致肌动蛋白积累减少,对NK细胞的易感性增强。
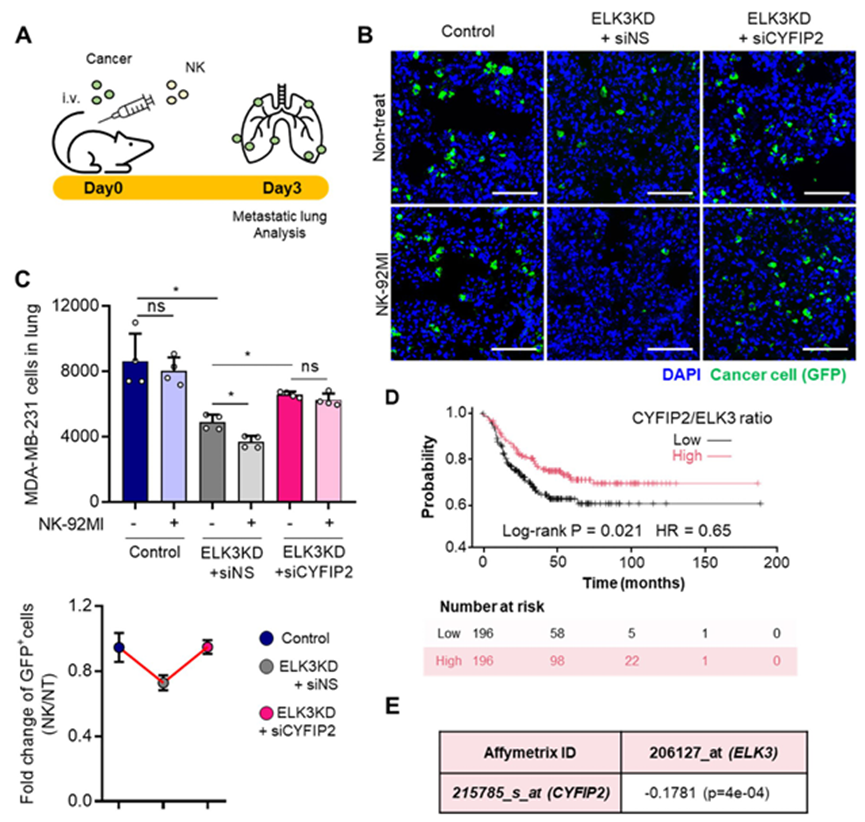
6. ELK3-CYFIP2轴在TNBC对NK细胞免疫监视的反应和TNBC患者的生存中发挥作用
为确定ELK3-CYFIP2轴是否调控TNBC的转移能力以及体内的NK反应,对注射GFP表达对照细胞、ELK3KD细胞或CYFIP2沉默的ELK3KD MDA-MB-231细胞的小鼠进行外渗分析。向免疫缺陷的 NSG 小鼠静脉注射总计 1×106 个癌细胞,1 小时后再注射 3×106 个 NK-92MI 细胞(图 6A)。3 天后检查肺部是否存在 GFP 阳性细胞。在各组小鼠肺组织的冰冻切片中观察到 GFP 阳性癌细胞(图 6B)。与体外实验结果一致,与对照组 MDA-MB-231 细胞相比,注射了 ELK3KD 细胞的小鼠检测到的 GFP 阳性细胞较少,而 CYFIP2 沉默的 ELK3KD 细胞检测到的 GFP 阳性细胞水平与对照组细胞相似。值得注意的是,当向已注射对照细胞或 CYFIP2 沉默的 ELK3KD 细胞的小鼠注射 NK-92MI 细胞时,GFP 阳性细胞的数量保持不变;但在注射 ELK3KD 细胞的小鼠中,GFP 阳性细胞的数量显著下降。通过流式细胞术对肺部的 GFP 阳性细胞进行量化(图 6C)。在每组小鼠的肺组织中都观察到一致的GFP阳性癌细胞,这表明ELK3-CYFIP2轴影响MDA-MB-231细胞对体内NK细胞抗癌活性的反应。
最后,使用KM Plotter在线工具研究ELK3和CYFIP2在TNBC细胞系和患者中表达的临床意义。根据CYFIP2/ELK3的比例,TNBC患者(n = 392)被分为 “高 ”和 “低 ”两组。Kaplan-Meier 图显示,“低 ”组患者的生存期明显短于 “高 ”组患者(图 6D)。此外,ELK3 的表达与 CYFIP2 的表达呈微弱但有统计学意义的负相关(图 6E)。综上所述,这些研究结果表明,ELK3-CYFIP2轴具有与TNBC患者生存相关的生物学活性。
结论
综上所述,本研究有力地支持这样一种观点,即ELK3-CYFIP2轴是驱动TNBC转移的肌动蛋白重塑调节因子,它同时影响癌细胞的迁移和对NK细胞的免疫敏感性,以ELK3-CYFIP2轴为靶点有望成为对抗转移性TNBC的治疗策略。具体来说,抑制该轴可以阻碍癌细胞的迁移和粘附,同时提高对NK细胞的免疫敏感性。破坏这一通路有望为 TNBC 患者制定更有效的治疗策略铺平道路。
实验方法
细胞培养,细胞转染,细胞迁移,细胞粘附试验,qRT-PCR,荧光素酶实验,Western blot,ChIP,CFSE/7-AAD 检测,免疫组化,延时观察癌细胞在 NK 细胞存在下的肌动蛋白堆积情况
参考文献
Choi SH, Jang HJ, Park JD, Ryu KS, Maeng E, Cho S, Park H, Jung HY, Park KS. ELK3-CYFIP2 axis-mediated actin remodeling modulates metastasis and natural killer cell responses in triple-negative breast cancer. J Exp Clin Cancer Res. 2025 Feb 10;44(1):48. doi: 10.1186/s13046-025-03309-7. PMID: 39930469; PMCID: PMC11808954.